
Abstract
Some recent studies have been previously suggested that nanoparticulate titanium dioxide (TiO2) damaged liver function and decreased immunity of mice, but the spleen injury and its oxidative stress mechanism are still unclear. To understand the spleen injury induced by intragastric administration of nanoparticulate anatase TiO2 for consecutive 30 days, the spleen pathological changes, the oxidative stress, and p38 and c-Jun N-terminal kinase signaling pathways, along with nuclear factor-κB and nuclear factor-E2-related factor-2 (Nrf-2), were investigated as the upstream events of oxidative stress in the mouse spleen from exposure to nanoparticulate TiO2. The results suggested that nanoparticulate TiO2 caused congestion and lymph nodule proliferation of spleen tissue, which might exert its toxicity through oxidative stress, as it caused significant increases in the mouse spleen reactive oxygen species accumulations, subsequently leading to the strong lipid peroxidation and the significant expression of heme oxygenase-1 via the p38-Nrf-2 signaling pathway. The studies on the mechanism by which nanoparticulate TiO2 induced the p38-Nrf-2 signaling pathway are helpful to a better understanding of the nanoparticulate TiO2-induced oxidative stress and reduction of immune capacity.
Similar content being viewed by others


Introduction
Nanoparticles and materials are being rapidly produced in large quantities throughout the world. Widespread application of nanomaterials confers enormous potential for human exposure and environmental release. Recently, however, scientists and organizations have raised the environmental and other safety concerns about nanotechnology [1–3]. As new types of photo-catalyst, anti-ultraviolet light agents, and photoelectric effect agents, titanium dioxide (TiO2) nanoparticles are used in a variety of consumer products (such as toothpastes, sunscreens, cosmetics, food products) [4], paints and surface coatings [5], and in the environmental decontamination of air, soil, and water [6, 7]. As an ultrafine-sized material, the nanoparticulate TiO2 can enter the human body through various routes such as inhalation (respiratory tract), ingestion (gastrointestinal tract), dermal penetration (skin), and injection (blood circulation) [8, 9].
Although little is known about nanoparticulate TiO2 toxicity, oxidative stress, which elicits a wide variety of cellular events, such as apoptosis, cell cycle arrest, and the induction of antioxidant enzymes, has often been reported as nanoparticulate TiO2- induced toxicity. Numerous previous studies on nanoparticulate TiO2 toxicity, with various animal organ types, such as lung, gill, brain, liver, and kidney, have reported that oxidative stress is one of the most important toxicity mechanisms related to the exposure to nanoparticulate TiO2 [10–14]. Nanoparticulate TiO2 was demonstrated to damage the haemostasis blood system and immune responses in mice [15]. The reduction of immune responses of mice caused by nanoparticulate anatase TiO2 may be due to the spleen damage. Spleen is the largest immune organ in humans, participating in immune response, generating lymphocytes, eliminating aging erythrocytes, and storing blood. However, we need to be focused on whether nanoparticulate TiO2 induces pathological changes to spleen and how nanoparticulate TiO2 damages spleen.
In order to further elucidate the molecular mechanism of nanoparticulate TiO2-induced oxidative stress in the mouse spleen, the evaluation in the toxicity needs to be focused on involving the oxidative stress responding signal transduction pathway and transcription factors caused by nanoparticulate TiO2. The studies of the upstream signaling mechanism responsible for regulating oxidative stress have been focused on the mitogen-activated protein (MAP) kinase cascades, including p38 and c-Jun N-terminal kinase (JNK) [16]. However, the upstream signaling mechanism responsible for regulating the oxidative stress involved in nanoparticulate TiO2 toxicity is rarely reported. The previous studies demonstrated that JNKs and p38 MAP kinases involving MAP kinase cascades are preferentially activated by various stresses, such as X-ray or UV irradiation, heat or osmotic shock, and oxidative or nitrosative stress [17–20]. Moreover, redox-sensitive transcription factors, such as nuclear factor kB (NF-κB) and nuclear factor-E2-related factor-2 (Nrf-2), can also be evaluated as target transcription factors of nanoparticulate TiO2 toxicities. NF-κB, activated by oxidative stress, induces the expression of a variety of proteins that function in the immunological and cellular detoxifying defense systems [21, 22] and has been identified as a transcription factor regulated by the intracellular redox status [23]. When activated by oxidative stress, Nrf-2 breaks free from Kelchlike ECH-associated protein 1 (Keap1) and translocates into the nucleus, where it binds to an antioxidant response factor, a cis-acting enhancer sequence that mediates the transcriptional activation of genes in response to oxidative stress, including heme oxygenase-1 (HO-1) [24]. Heme oxygenases (HO) are rate-limiting enzymes that catalyze the conversion of heme into carbon monoxide and biliverdin [25]. They have antioxidant capacity and therefore act as potent anti-inflammatory proteins whenever oxidative injury takes place [26]. HO-2 is constitutively produced within the brain and testes, whereas HO-1 is produced ubiquitously, but only marginally in the resting state [27]. Rapid induction of HO-1 follows various stresses [28–30]. The previous studies have suggested protective roles of HO-1 in various inflammatory conditions [31, 32]. Many studies have suggested that the transcription factor Nrf-2 plays an essential role in the antioxidant response factor-mediated expression of phase II detoxifying and antioxidant enzymes, as well as other stress-inducible genes, in response to oxidative stress [33–38].
In this article, the spleen pathological changes, the oxidative stress, the expression levels of the oxidative stress genes and their proteins including p38, JNK, NF-κB, Nrf-2, and HO-1 in the mouse spleen were investigated to further understand mechanism of the splenic injury in mice caused by nanoparticulate TiO2.
Materials and Methods
Chemicals and Preparation
TiO2 (100% anatase, CAS #:13463-67-7, VK-TA05) was purchased from Hangzhou Wanjing New Material Co. Ltd. (Hangzhou, China) (the particle characteristics are shown in Table 1) and the particles were used in this experiment.
A 0.5% hydroxypropyl-methylcellulose K4M (HPMC, K4M) was used as a suspending agent. Each TiO2 powder was dispersed onto the surface of 0.5%, w/v HPMC solution, and then the suspending solutions containing TiO2 particles were treated by ultrasonic for 15–20 min and mechanically vibrated for 2 or 3 min.
Animals and Treatment
Eighty CD-1 (ICR) female mice (22 ± 2 g) were purchased from the Animal Center of Soochow University. Animals were housed in stainless steel cages in a ventilated animal room. Room temperature was maintained at 20 ± 2°C, relative humidity at 60 ± 10%, and a 12-h light/dark cycle. Distilled water and sterilized food for mice were available ad libitum. They were acclimated to this environment for 5 days prior to dosing. All animal procedures were performed in compliance with the regulations and guidelines of the international ethics committee on animal welfare. Animals were randomly divided into four groups: control group (treated with 0.5% HPMC) and three experimental groups (5, 50, and 150 mg/kg BW nano-anatase TiO2). Nanoparticulate anatase TiO2 (5, 50, and 150 mg/kg BW) suspensions were given to mice by intragastric administration every day for 30 days, respectively. The control group was treated with 0.5% HPMC. The symptom and mortality were observed and recorded carefully everyday for 30 days. After 30 days, all animals were first weighed and then sacrificed after being anesthetized by ether. The spleens were excised and weighed.
Histopathological Examination of Spleen
Histological observations were performed according to the standard laboratory procedures. Mice (four mice/control and three treatment groups) at the end of day 30 were dissected for histology. A small piece of spleen, fixed in 10% (v/v) formalin, was embedded in a paraffin block, sliced into 5 μm thicknesses and then placed onto glass slides. The section was stained with hematoxylin–eosin (HE) and examined by optica microscopy (Nikon U-III Multi-point Sensor System: Biodirect-Inc., Nikon, USA), and the identity and analysis of the pathology slides were blind to the pathologist.
ROS and Lipid Peroxidation Assay
ROS Assay
Superoxide ion (O ·–2 ) in spleen tissue was measured as described previously by Oliveira et al. [39], by determining the reduction of 3′-[1-[phenylamino-carbonyl]-3,4- tetrzolium]-bis (4-methoxy-6-nitro) benzenesulfonic acid hydrate (XTT) in the presence of O ·–2 , with some modifications. The spleen was homogenized with 2 mL of 50 mM Tris–HCl buffer (pH 7.5) and centrifuged at 5,000×g for 10 min. The reaction mixture (1 mL) contained 50 mM Tris–HCl buffer (pH 7.5), 20 μg spleen proteins, and 0.5 mM XTT. The reaction of XTT was determined at 470 nm for 5 min. Corrections were made for the background absorbance in the presence of 50 units of superoxide dismutase (SOD). The production rate of O ·–2 was calculated using an extinction coefficient of 2.16 × 104 M–1 cm–1.
The detection of H2O2 contents in the liver tissues was carried out by the xylenol orange assay [40], with minor modifications. In short, after the preincubation of P2 with mercurials and/or quercetin/catalase (30 min at 25°C), the reaction medium was centrifuged at 17,500×g for 10 min at 4°C and the supernatant was incubated for 30 min in a reaction medium containing 250 mM perchloric acid, 2.5 mM ammonium iron (II) sulfate hexahydrate, and 1 mM xylenol orange. Hydroperoxide levels were determined at 560 nm using a hydrogen peroxide curve as standard.
Lipid Peroxidation
Spleen lipid peroxidation was determined as the concentration of malondialdehyde (MDA) generated by the thiobarbituric acid (TBA) reaction as described by Buege and Aust [41], but with the introduction of an isobutanol extraction step for the removal of interfering compounds. For analysis, a subsample of tissue was thawed, homogenized, and cells lysed using a 4% TBA solution in 0.2M HCl. The reaction mixture was then incubated at 90°C for 45 min. The resulting TBA–MDA adduct was phase-extracted using isobutanol. The isobutanol phase was then read at a wavelength of 535 nm on a UV-3010 spectrophotometer. MDA standard curves were prepared by acid hydrolysis of 1,1,3,3-tetramethoxypropane.
Expression Assay of Oxidative Stress Genes and Proteins
The mRNA expression of regulating the oxidative stress genes, including p38, JNK, NFR2, NF-κB, and HO-1, were determined by real-time quantitative reverse transcriptase–polymerase chain reaction (RT-PCR) [42–44]. Spleens in the same growth period from the three different treatments were used. The right spleens from mice with or without nano-anatase TiO2 treatment were homogenized using QIAzol lysis reagent with a Tissue Ruptor (Roche, Indianapolis, IN). Total RNA from the homogenates was isolated using Tripure Isolation Reagent (Roche) according to the manufacturer’s instructions. The RT reagent (ShineGene: Shanghai Shinegene Co., China) of 30 μL was prepared by mixing 15 μL of 2 × RT buffer, 1 μL random primer in a concentration of 100 pmol·μL−1, 1 μL of RTase, 5 μL RNA, and 8 μL DEPC water together. The reaction condition was 25°C for 10 min, 40°C for 60 min, and 70°C for 10 min.
Synthesized cDNA was used for the real-time PCR. Primers were designed using Primer Express Software according to the software guidelines (Table 2).
All primers were purchased from ShineGene. For the 50-μL PCR reaction, 25 μL 2× PCR buffer, 0.6 μL 2× primers (25 pmol·μL−1), 0.3 μL probe (25 pmol·μL−1), 1 μL cDNA, and 22.8 μL DEPC water (Sigma) were mixed together. The parameters for a two-step PCR were 94°C for 3 min, 94°C for 20 s, 60°C for 20 s, then 72°C for 20 s, 35 cycles.
The gene expression analysis and experimental system evaluation were performed according to the standard curve and quantitation reports.
To determine p38, JNK, NF-κB, Nfr2, and HO-1 levels of the spleen, enzyme-linked immunosorbent assay (ELISA) was performed by using commercial kits that are selective for the mouse spleen p38, JNK, NF-κB, Nfr2, and HO-1 (RD Systems, Minneapolis, USA). The manufacturer’s instruction was followed. The absorbance was measured on a microplate reader at 450 nm (Varioskan Flash, Thermo Electron, Finland), and the p38, JNK, NF-κB, Nfr2, and HO-1 concentration of samples was calculated from a standard curve.
Statistical Analysis
Statistical analyses were done using SPSS11.5 software. Data were expressed as means ± SE. One-way analysis of variance was carried out to compare the differences of means among multigroup data. Dunnett’s test was carried out when each group of experimental data was compared with solvent-control data. Statistical significance for all tests was judged at a probability level of 0.05.
Result
Histopathological Evaluation
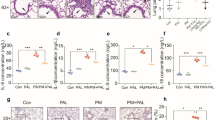
The histological photomicrographs of the spleen sections are shown in Fig. 1. No severe damages of spleen tissue were reflected in the 5 mg/kg nanoparticulate anatase TiO2-treated group (Fig. 1b) compared to the control (Fig. 1a), while the congestion of the spleen tissue was showed in the 50 mg/kg nanoparticulate anatase TiO2 group (Fig. 1c) and lymph nodule proliferation was observed in the 150 mg/kg nanoparticulate anatase TiO2-treated group (Fig. 1d). We observed histological photomicrographs of the spleens of four mice of each group, indicating that the 50 and 150 mg/kg nanoparticulate anatase TiO2 caused histological changes in the same manner.

Histopathological observation of spleen caused by intragastric administration with nanoparticulate anatase TiO2 for 30 days (HE, ×original magnification). a Control group (×100) presents no abnormal pathology; b 5 mg/kg nanoparticulate TiO2 group (×200) presents no abnormal pathology; c 50 mg/kg nanoparticulate TiO2 group (×200) presents congestion (arrow); d 150 mg/kg nanoparticulate group (×100) presents lymph nodule proliferation (arrows)
ROS Production and Lipid Peroxidation
The effects of treatments with various doses of nanoparticulate TiO2 on the accumulation of O2 · −, H2O2, and the MDA content in the mouse spleen are shown in Figs. 2 and 3, respectively. The significant differences were observed the production rate of ROS and the MDA content of the spleen caused by nanoparticulate TiO2 (p < 0.05 or 0.01), e.g. O · −2 was increased by 29.38%, 75.41%, and 145.01%, respectively; H2O2 was promoted by 25.82%, 57.84%, and 109.53%, respectively; MDA was enhanced by 29.36%, 63.65%, and 191.19%, respectively. This phenomenon showed that higher-dose nanoparticulate TiO2 promoted the generation of oxidative stress and caused lipid peroxidation in the mouse spleen.

The ROS accumulation in the mouse spleen after intragastric administration with nanoparticulate TiO2 for 30 days. Bars marked with an asterisk or double asterisk indicate significant difference from the control (no nanoparticules) at the 5% or 1% confidence level, respectively. Values represent means ± SE, n = 5

The MDA content of spleen of ICR mice after intragastric administration with nanoparticulate TiO2 for 30 days. Bars marked with double asterisk indicate significant difference from the control (no nanoparticules) at the 1% confidence level. Values represent means ± SE, n = 5
MAP Kinase Signal Transduction and Transcription Factors
The damage happened in the spleen according to the histopathological observations. To confirm expression levels of the oxidative stress genes and their proteins including p38, JNK, NF-κB, Nrf-2, and HO-1 in the nanoparticulate TiO2-induced spleen injury, real-time quantitative RT-PCR and ELISA were used to demonstrate the changes of the oxidative stress genes and their proteins levels in the nanoparticulate TiO2-treated mice (Tables 3 and 4). In Tables 3 and 4, we observed that nanoparticulate TiO2 induced a dramatic increase of p38, JNK, NF-κB, Nrf-2, and HO-1 expression in the treated mouse spleen (P < 0.05 or 0.01). The extents of increase of these genes and their proteins are consistent with the trends on the histological photomicrograph of the spleen sections in the exposed mice.
Discussion
Wang et al. [45] observed that the coefficients of the mouse spleen increased only a little, and there are no abnormal pathology changes in the spleen after a fixed large dose of 5 g/kg BW of nanoparticulate TiO2 (25 and 80 nm) was administered by a single oral gavage for 2 weeks. However, Chen et al., by histopathological examinations, observed that nano-TiO2 particles caused a severe lesion of spleen and a mass of neutrophilic cells in spleen tissues by intraperitoneal injection, revealing that inflammation in spleen tissues was very serious [46]. Our researches also observed that the coefficients of the spleen of mice significantly increased by intragastric administration or intraperitoneal injection of higher doses of nanoparticulate TiO2 (5 nm) for 30 or 14 days [15, 47]. The results of this study indicate that the intragastric administration of higher doses of nanoparticulate anatase TiO2 can induce histopathological changes of spleen, including the congestion and lymph nodule proliferation, suggesting an inflammation in spleen tissues (Fig. 1). The present study indicates that the spleen lesion of mice is triggered by nanoparticulate TiO2 oxidative stress generation and activation of the oxidative stress genes that resulted in inflammation of spleen tissue.
To prove spleen oxidative stress of mice, we detected ROS and MDA contents. The studies showed that the obvious production of ROS (such as O .–2 and H2O2) and lipid peroxidation (MDA content increased) occurred in the mouse spleen treated with higher nanoparticulate anatase TiO2 doses (Figs. 2 and 3), indicating that these nanoparticulate anatase TiO2-treated mouse spleen underwent severe oxidative stress. Similarly, nanoparticulate anatase TiO2 was reported to cause oxidative stress in the mouse brain, liver and kidney [12–14]. It has been demonstrated that nanoparticules are mediating their toxicity through production of ROS and that the level of ROS depends on the chemistry and structure of the nanoparticules [4, 9, 10, 48, 49]. The overproduction of ROS would break down the balance of the oxidative/antioxidative system in the spleen, resulting in the lipid peroxidation, which is closely related to the reduction of the antioxidative capacity. In our previous studies, nanoparticulate anatase TiO2 inhibited the activities of SOD, catalase, ascorbic acid peroxidase, and glutathione peroxidase, and decreased ascorbic acid and reduced glutathione in the mouse brain, liver, and kidney [12–14]. In fact, any chemical damage is based on the physical binding interaction. Some intermolecular action mechanisms were also demonstrated to be related to the oxidative stress [50, 51].
To further understand the mechanism of oxidative stress due to exposure to nanoparticulate TiO2, the detection of molecular events, such as MAP kinase signal transduction and subsequent transcription factor activation (Tables 3 and 4), was undertaken. The expressions of p38 and JNK were significantly induced by nanoparticulate TiO2. The overall results for ROS formation and on MAP kinase signaling studies suggested that nanoparticulate TiO2 provokes oxidative stress, and in response to this, mainly the p38 and JNK MAP kinase signaling pathway seems to be activated. As a downstream event of MAP kinase signaling, NF-κB and Nrf-2 transcription factors, which are known to respond to oxidative stress, were examined in the mouse spleen treated with nanoparticulate TiO2 (Tables 3 and 4). Nanoparticulate TiO2 induced the expressions of NF-κB and Nrf-2. To further investigate the cellular consequences of oxidative stress signaling through p38-Nrf-2 activation by nanoparticulate TiO2 exposure, representative antioxidant enzymes, such as HO-1, were examined (Tables 3 and 4). HO-1 is a relatively novel enzyme, with potent anti-inflammatory and cytoprotective antioxidant effects [52–55]. The expression of HO-1 protein was dramatically increased in nanoparticulate TiO2-treated mice. The induction of HO-1 due to exposure to nanoparticulate TiO2 may be mediated through p38 MAP kinase and the Nrf-2 signal transduction pathway. Recently, Lim et al. reported that the cyclopentenone prostaglandin compound stimulates HO-1 expression through the p38 MAP kinase and Nrf-2 pathway in rat vascular smooth muscle cells [56, 57]. Induction of HO-1 can be interpreted as a cellular defense mechanism against oxidative stress; it is well known that HO-1 induction is regulated by Nrf-2 activation [24]. In this study, nanoparticulate TiO2-induced NF-κB activation was not observed, which was unexpected, as NF-κB is the major stress response transcription factor and has been reported to respond to a wide variety of environment stressors.
Conclusion
Overall, the results of experimental studies suggest that nanoparticulate TiO2 may exert their toxicity through oxidative stress. Nanoparticulate TiO2 causes congestion and lymph nodule proliferation of spleen tissue of mice and a significant increase in the spleen ROS productions, and subsequently leads to a strong induction of HO-1 via the p38-Nrf-2 signaling pathway. The tested oxidative stress parameters in this study were rather limited in terms of allowing a full understanding of the oxidative stress and spleen response due to exposure to nanoparticulate TiO2. Further studies on the mechanism by which nanoparticulate TiO2 induce the p38-Nrf-2 signaling pathway to better understand the nanoparticulate TiO2-induced oxidative stress, as well as with concentration– response and time-course analyses, are warranted.
References
Dagani R (2003) Nanomaterials: safe or unsafe? Chem Eng News 81:30–33
Service RF (2003) American Chemical Society meeting. Nanomaterials show signs of toxicity. Science 300:243
Warheit DB (2004) Nanoparticles: health impacts? Mater Today 7:32–35
Gurr J, Wang ASS, Chen C, Jan K (2005) Ultrafine titanium dioxide particle in the absence of photoactivation can induce oxidative damage to human bronchial epithelial cells. Toxicol 213:66–73
J Fisher, T Egerton (2001) Kirk-Othmer encyclopedia of chemical technology. New York: John Wiley & Sons, Titanium Compounds, Inorganic
Choi H, Stathatos E, Dionysiou DD (2006) Solgel preparation of mesoporous photocatalytic TiO2 films and TiO2/Al2O3 composite membranes for environmental applications. Appl Catal B-Environ 63:60–67
Esterkin CR, Negro AC, Alfano OM, Cassano AE (2005) Air pollution remediation in a fixed bed photocatalytic reactor coated with TiO2. AIChE J 51:2298–2310
Jin CY, Zhu BS, Wang XF, Lu QH (2008) Cytotoxicity of titanium dioxide nanoparticles in mouse fibroblast cells. Chem Res Toxicol 21(9):1871–1877
Oberdörster G, Oberdörster E, Oberdörster J (2005) Nanotoxicology: an emerging discipline evolving from studies of ultrafine particles. Environ Health Perspect 113(7):823–839
Afaq F, Abidi P, Matin R, Rahman Q (1998) Cytotoxicity, pro-oxidant effects and antioxidant depletion in rat lung alveolar macrophages exposed to ultrafine titanium dioxide. J Appl Toxicol 18:307–312
Federici G, Shaw BJ, Handy RD (2007) Toxicity of titanium dioxide nanoparticles to rainbow trout (Oncorhynchus mykiss): gill injury, oxidative stress, and other physiological effects. Aquat Toxicol 84:415–430
Ma LL, Liu J, Li N, Wang J, Duan YM, Yan JY, Liu HT, Wang H, Hong FS (2010) Oxidative stress in the brain of mice caused by translocated nanoparticulate TiO2 delivered to the abdominal cavity. Biomaterials 31:99–105
Liu HT, Ma LL, Liu J, Zhao JF, Yan JY, Hong FS (2010) Toxicity of nano-anatase TiO2 to mice: liver injury, oxidative stress. Toxicol Environ Chem 92(1):175–186
Zhao JF, Wang J, Wang SS, Zhao XY, Yan JY, Ruan J, Li N, Duan YM, Wang H, Hong FS (2010) The mechanism of oxidative damage in nephrotoxicity of mice caused by nano-anatase TiO2. J Exp Nanoscience doi:10.1080/17458081003628931, 2010 in press
Duan YM, Liu J, Ma LL, Li N, Liu HT, Wang J, Zheng L, Liu C, Wang XF, Zhang XG, Yan JY, Wang H, Hong FS (2010) Toxicological characteristics of nanoparticulate anatase titanium dioxide in mice. Biomaterials 31:894–899
Kyriakis JM, Avruch J (2001) Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol Rev 81:807–869
Takeda K, Matsuzawa A, Nishitoh H, Ichijo H (2003) Roles of MAPKKK ASK1 in stress-induced cell death. Cell Struct Funct 28:23–29
Hagemann C, Blank JL (2001) The ups and downs of MEK kinase interactions. Cell Signall 13(12):863–875
Qadri I, Iwahashi M, Capasso JM, Hopken MW, Flores S, Schaack J, Simon FR (2004) Induced oxidative stress and activated expression of manganese superoxide dismutase during hepatitis C virus replication: role of JNK, p38 MAPK and AP-1. Biochem J 378:919–928
Camacho-Barquero L, Villegas I, Sánchez-Calvo JM, Talero E, Sánchez-Fidalgo S, Motilva V, Alarcón de la Lastra C (2007) Curcumin, a Curcuma longa constituent, acts on MAPK p38 pathway modulating COX-2 and iNOS expression in chronic experimental colitis. Int Immunopharmacol 7:333–342
Janssen YM, Barchowsky A, Treadwell M, Driscoll KE, Mossman BT (1995) Asbestos induces nuclear factor kappa B (NF-kappa B) DNA-binding activity and NF-kappa B-dependent gene expression in tracheal epithelial cells. Proc Natl Acad Sci USA 92:8458–8462
Pinkus R, Weiner LM, Daniel V (1996) Role of oxidants and antioxidants in the induction of AP-1, NF-kappaB, and glutathione S-transferase gene expression. J Biol Chem 271:13422–13429
Sen CK, Packer L (1996) Antioxidant and redox regulation of gene transcription. FASEB J 10:709–720
Itoh K, Wakabayashi N, Katoh Y, Ishii T, O’connor T, Yamamoto M (2003) Keap1 regulates both cytoplasmic-nuclear shuttling and degradation of Nrf2 in response to electrophiles. Genes Cells 8:379–391
Maines MD (1988) Heme oxygenase: function, multiplicity, regulatory mechanism, and clinical applications. FASEB J 2:2557–2568
Choi AM, Alam J (1996) Heme oxygenase-1: function, regulation, and implication of a novel stress-inducible protein in oxidant-induced lung injury. Am J Respir Cell Mol Biol 15(1):9–19
Maines MD (1997) The heme oxygenase system: a regulator of second messenger gases. Annu Rev Pharmacol Toxicol 37:517–554
Okinaga S, Takahashi K, Takeda K, Yoshizawa M, Fujita H, Sasaki H, Shibahara S (1996) Regulation of human heme oxygenase-1 gene expression under thermal stress. Blood 87:5074–5084
Cantoni L, Rossi C, Rizzardini M, Gardina M, Ghezzi P (1991) Interleukin-1 and tumor necrosis factor induce hepatic haeme oxygenase. Biochem J 279:891–894
Chen K, Gunter K, Maines MD (2002) Neurons overexpressing heme oxygenase-1 resist oxidative stress-mediated cell death. J Neurochem 75(1):304–313
Willis D, Moore AR, Frederick R, Willoughby DA (1996) Heme oxygenase: a novel target for the modulation of the inflammatory response. Nat Med 2:87–90
Yachie A, Niida Y, Wada T, Igarashi N, Kaneda H, Toma T, Ohta K, Kasahara Y, Koizumi S (1999) Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1 deficiency. Clin Invest 103(1):129–135
Itoh K, Chiba T, Takahashi S, Ishii T, Igarashi K, Katoh Y, Oyake T, Hayashi N, Satoh K, Hatayama I, Yamamoto M, Nabeshima Y (1997) An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem Biophy Res Commun 236:313–322
Chan K, Kan YW (1999) Nrf2 is essential for protection against acute pulmonary injury in mice. Proc Natl Acad Sci USA 96:12731–12736
Hayes JD, Chanas SA, Henderson CJ, McMahon M, Sun C, Moffat GJ, Wolf CR, Yamamoto M (2000) The Nrf2 transcription factor contributes both to the basal expression of glutathione S-transferases in mouse liver and to their induction by the chemopreventive synthetic antioxidants, butylated hydroxyanisole and ethoxyquin. Biochem Soc Trans 28:33–41
Chan K, Han XD, Kan YW (2001) An important function of Nrf2 in combating oxidative stress: detoxification of acetaminophen. Proc Natl Acad Sci USA 98:4611–4616
Kim YC, Masutani H, Yamaguchi Y, Itoh K, Yamamoto M, Yodoi J (2110) Hemin induced activation of the thioredoxin gene by Nrf2. A differential regulation of the antioxidant responsive element by a switch of its binding factors. J Biol Chem 276:18399–18406
Kwak MK, Itoh K, Yamamoto M, Kensler TW (2002) Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: role of antioxidant response element-like sequences in the nrf2 promoter. Mol Cell Biol 22:2883–2892
Oliveira CP, Lopasso FP, Laurindo FR, Leitao RM, Laudanna A (2001) Protection against liver ischemia-reperfusion injury in rats by silymarin or verapamil. Transplant Proc 33:3010–3014
Nourooz-Zadeh J, Tajaddini-Sarmadi J, Wolff SP (1994) Measurement of plasma hydroperoxide concentrations by the ferrous oxidation-xylenol orange assay in conjunction with triphenylphosphine. Anal Biochem 220:403–409
Buege JA, Aust SD (1978) Microsomal lipid peroxidation. Meth Enzymol 52:302–310
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25:402–408
Ke LD, Chen Z (2000) A reliability test of standard-based quantitative PCR: exogenous vs endogenous standards. Mol Cell Probes 14(2):127–135
Liu WH, David Saint A (2002) Validation of a quantitative method for real time PCR kinetics. Biochem Biophys Res Commun 294:347–353
Wang JX, Zhou GQ, Chen CY, Yu HW, Wang TC, Ma YM, Jia G, Gao YX, Li B, Sun J, Li YF, Jia F, Zhao YL, Chai ZF (2007) Acute toxicity and biodistribution of different sized titanium dioxide particles in mice after oral administration. Toxicol Lett 168:176–185
Chen JY, Dong X, Zhao J, Tang GP (2009) In vivo acute toxicity of titanium dioxide nanoparticles to mice after intraperitioneal injection. J Appl Toxicol 29:330–337
Liu HT, Ma LL, Zhao JF, Liu J, Yan JY, Ruan J, Hong FS (2009) Biochemical toxicity of nano-anatase TiO2 particles in mice. Biol Trace Elem Res 129(1):170–180
Long TC, Saleh N, Tilton RD, Lowry G, Veronesi B (2006) Titanium dioxide (P25) produces reactive oxygen species in immortalized brain microglia (BV2): implications for nanoparticle neurotoxicity. Environ Sci Technol 40:4346–4352
Long TC, Tajuba J, Sama P, Sale N, Swartz C, Parker J, Hester S, Lowry GV, Veronesi B (2007) Nano-TiO2 stimulates ROS in brain microglia and damages neurons in vitro. Environ Health Perspect 115:1631–1637
Gao HW, Xu Q, Chen L, Wang SL, Wang Y, Wu LL, Yuan Y (2008) Potential protein toxicity of synthetic pigments, binding of poncean S to human serum albumin. Biophys J 94:906–917
Xu Z, Liu XW, Ma YS, Gao HW (2010) Interactions of nano-TiO2 with lysozyme: insights into the enzyme toxicity of nanosized particles. Environ Sci Pollut Res 17:798–806
Maines MD (1988) Hemeoxygenase: function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J 2:2557–2568
Morse D, Choi AM (2002) Heme oxygenase-1: the “emerging molecule” has arrived. Am J Respir Cell Mol Biol 27:8–16
Otterbein LE, Soares MP, Yamashita K, Bach FH (2003) Heme oxygenase-1: unleashing the protective properties of heme. Trends Immunol 24:449–455
Chen CY, Jang JH, Li MH, Surh YJ (2005) Resveratrol upregulates heme oxygenase-1 expression via activation of NF-E2-related factor 2 in PC12 cells. Biochem Biophys Res Commun 331:993–1000
Lee JS, Surh YJ (2005) Nrf2 as a novel molecular target for chemoprevention. Cancer Lett 224:171–184
Lim HJ, Lee KS, Lee S, Park JH, Choi HE, Go SH, Kwak HJ, Park HY (2007) 15d-PGJ2 stimulates HO-1 expression through p38 MAP kinase and Nrf-2 pathway in rat vascular smooth muscle cells. Toxicol Appl Pharmacol 223:20–27
Acknowledgements
This work was supported by the National Natural Science Foundation of China (grant no. 30901218), the Medical Development Foundation of Soochow University (grant no. EE120701, China), the National Bringing New Ideas Foundation of Student of China (grant nos. 57315427, 57315927), and the Bringing New Ideas Foundation of Postgraduate of Medical College of Soochow University (China) and the Soochow University Start-up Fund (grant no. Q4134918, China).
Author information
Authors and Affiliations
Corresponding author
Additional information
Jue Wang, Na Li, Lei Zheng and Ying Wang contributed equally to this work.
Rights and permissions
About this article
Cite this article
Wang, J., Li, N., Zheng, L. et al. P38-Nrf-2 Signaling Pathway of Oxidative Stress in Mice Caused by Nanoparticulate TiO2 . Biol Trace Elem Res 140, 186–197 (2011). https://doi.org/10.1007/s12011-010-8687-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12011-010-8687-0