
This ancient and hostile ecosystem hosts a surprising variety of microbial organisms.
Abstract
The Rio Tinto, known by the Phoenicians as 'Ur-yero', or 'River of Fire', because of its deep red colour and high acidity, flows through the world's largest pyritic belt in southwestern Spain. Surprisingly, eukaryotic microbes are the principal contributors of biomass in this hostile river, which has a pH of 2 and contains much higher concentrations of heavy metals than are typically found in fresh waters. Here we show that the Rio Tinto shows an unexpected degree of eukaryotic diversity and includes new lineages that we have identified by sequence analysis of genes encoding small-subunit ribosomal RNAs. The diversity of these eukaryotes is much greater than that of prokaryotes, whose metabolism is responsible for the extreme environment.
Similar content being viewed by others

Main
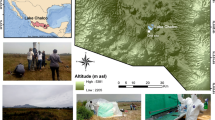
The Rio Tinto (Fig. 1) is 90 km long. Unlike other extreme acidic environments, the low pH and high concentrations of heavy metals in this vast ecosystem (3–20 g iron per litre of water, for example) predate 5,000 years of human mining activity, with iron-oxidizing bacteria having existed in the river basin some 300,000 years ago1.

J. L. DE LOPE & J. M. SANCHEZ
The river's red colour is due to the high concentration of iron dissolved in its extremely acidic waters.
Morphology-based investigations first alerted us to the possible variety of eukaryotic lineages in the Rio Tinto. Our molecular analysis reveals the extent of this eukaryotic diversity — which is quite unexpected in such an acidic and highly metal-rich environment — as well as several taxa and cosmopolitan organisms that have never previously been reported in extreme habitats (Fig. 2).

Bold letters indicate environmental clones. RT, sequences from Rio Tinto species; cul, sequences from cultured species. Underlined taxa represent genera that have been identified in the river by microscopy. Sampling sites: RT1, La Palma; RT3, Berrocal Upper; RT5i, the Origin, black filamentous biofilm; RT5ii, the Origin, green filamentous biofilm; RT7i, Anabel's Garden, green biofilm; RT7ii, Anabel's Garden, yellow biofilm. Bootstrap values are shown next to branches. Sequences have been deposited at GenBank under accession numbers AY082969–AY083001.
Eukaryotic algae contribute at least 60% of the biomass in the Rio Tinto2, forming conspicuous patches on the river's surface. Our molecular analysis detected many further, non-photosynthetic lineages, such as ciliates, cercomonads, vahlkampfiid amoebae, stramenopiles and fungi, which have all previously escaped detection3.
The phylogenetic placement of several sequences was consistent with the idea that they represent new eukaryotic lineages. Two groups of clones branched as possible sister taxa to amoeboid lineages4,5: the first of these (clones RT5iin14 and RT5iin16) branched at the base of the animal–fungal–nucleariid radiation; the second (RT5iin21 and RT5iin44) branched with the filose amoeba (Filamoeba nolandi), with moderately strong bootstrap support (83%). However, the long-branched nature of these sequences makes it difficult to position them precisely in a large phylogeny. The clone RT5iin25 may represent a new stramenopile lineage, with only weak bootstrap values supporting its relationship to Cafeteria roenbergensis.
The evolutionary distances between acidophilic species that occur in the river and their neutrophilic relatives offer insight into adaptation to an extremely acidic environment. We detected clones from the Rio Tinto that are closely related to neutrophilic species such as Chlamydomonas noctigama, Chlorella minutissima and Colpidium campylum. Adaptations associated with the transition from a neutral to an acidic environment must occur relatively rapidly when measured on evolutionary timescales.
We know from microscopic observations that rotifers also inhabit the river, as do heliozoa and other types of amoeba. The wide eukaryotic diversity associated with low pH and high concentrations of heavy metals indicates that adaptation to such adverse conditions must be more easily accomplished than was previously thought. As eukaryotes rarely flourish in extreme environments, our results challenge former ideas about the phylogenetic range of organisms that are capable of living in extreme environments.
References
Gónzalez-Toril, E. et al. in Biohydrometallurgy: Fundamentals, Technology and Sustainable Development (eds Ciminelli, V. S. T. & García, O.) 639–650 (Elsevier, Amsterdam, 2001).
Lopez-Archilla, A. I., Marin, I. & Amils, R. Geomicrobiol. J. 11, 223–233 (1994).
Durán, C., Marín, I. & Amils, R. in Biohydrometallurgy and the Environment: Towards the Mining of the 21st Century (eds Amils, R. & Ballester, A.) 521–530 (Elsevier, Amsterdam, 1999).
Amaral Zettler, L. A., Nerad, T. A., O'Kelly, C. J. & Sogin, M. L. J. Eukaryot. Microbiol. 48, 293–297 (2001).
Amaral Zettler, L. A. et al. Protist 151, 275–282 (2000).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Amaral Zettler, L., Gómez, F., Zettler, E. et al. Eukaryotic diversity in Spain's River of Fire. Nature 417, 137 (2002). https://doi.org/10.1038/417137a
Issue Date:
DOI: https://doi.org/10.1038/417137a
This article is cited by
-
The effect of type and combination of fertilizers on eukaryotic microbiome of date palm rhizosphere
Plant Growth Regulation (2024)
-
Extreme environments offer an unprecedented opportunity to understand microbial eukaryotic ecology, evolution, and genome biology
Nature Communications (2023)
-
New insights on the evolutionary relationships between the major lineages of Amoebozoa
Scientific Reports (2022)
-
Biogeochemical indicators (waters/diatoms) of acid mine drainage pollution in the Odiel river (Iberian Pyritic Belt, SW Spain)
Environmental Science and Pollution Research (2022)
-
Molecular diversity and biogeography of benthic microeukaryotes in temperate seagrass (Zostera japonica) systems of northern China
Acta Oceanologica Sinica (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
