
Abstract
Both clinical and preclinical research have shown that stress can potentiate drug use; however, the underlying mechanisms of this interaction are unknown. Previously, we have shown that a single exposure to forced swim (FS) reinstates extinguished conditioned place preference (CPP) to cocaine and that cAMP response element binding protein (CREB) is necessary for this response. CREB can be activated by corticotropin releasing factor (CRF) receptor type 1 (CRFR1) binding, which mediates neuroendocrine and behavioral responses to stress as well as to drugs of abuse. The present experiments investigate whether changes in cocaine reward elicited by previous exposure to stress are mediated by CREB and/or CRFR1. Chronic exposure to FS in advance of conditioning enhances cocaine CPP in wild-type mice, but this is blocked in CREB-deficient mice. In addition, pretreatment with the CRFR1 antagonist, antalarmin, before FS exposure blocks this stress-induced enhancement of cocaine CPP. Furthermore, FS-induced increase in phosphorylated CREB (pCREB), specifically in the lateral septum (LS) and nucleus accumbens (NAc) is also blocked by antalarmin. Taken together, these studies suggest that both CREB and CRFR1 activation are necessary for stress-induced potentiation of drug reward.
Similar content being viewed by others


INTRODUCTION
Prior exposure to stress can increase drug-taking behavior. In clinical studies, increased stress levels together with lack of support and coping skills are linked to increased cocaine, nicotine, alcohol, and marijuana use (Kaplan et al, 1986; Koob and Kreek, 2007; Maddahian et al, 1988a, 1988b; Sinha, 2007). In animals, acute exposure to stressors facilitates self-administration and conditioned place preference (CPP) to cocaine (Goeders and Guerin, 1994; Hadaway et al, 1979; Haney et al, 1995; McLaughlin et al, 2003; Miczek and Mutschler, 1996; Piazza et al, 1990; Piazza and Le Moal, 1998; Ramsey and Van Ree, 1993; Shaham and Stewart, 1994b). However, few studies have evaluated the effects of chronic stressors on drug-associated behaviors. Moreover, the molecular mechanisms associated with the ability of stress to facilitate drug reward have not been characterized. Previous work in our laboratory has demonstrated that the transcription factor cAMP response element binding protein (CREB) is critically involved in the ability of stress to reactivate cocaine CPP (Kreibich and Blendy, 2004). Therefore, we hypothesized that chronic forced swim stress (FS) would facilitate cocaine CPP and that this effect may also be mediated by CREB.
CREB plays a key role in the mechanisms underlying drug reward (Carlezon et al, 1998; Kreibich and Blendy, 2004; Valverde et al, 2004; Walters and Blendy, 2001; Walters et al, 2005); however, the signaling pathways through which CREB may be modulating interactions between stress and drug reward are unclear. The CRF system is important in responses to both stress and drugs of abuse (Sarnyai et al, 2001). Specifically, CRF receptor type 1 (CRFR1) mediates neuroendocrine and behavioral responses to stress as well as to drugs of abuse. The activation of CRFR1 stimulates the G protein Gαs, leading to the activation of protein kinase A, and the transcription factor CREB (Kasagi et al, 2002; McEvoy et al, 2002). As chronic stress has been shown to increase pCREB expression (Kwon et al, 2006), examining the role of CRFR1 activation in the ability of FS to induce phosphorylation of CREB could help delineate the signaling pathways underlying stress-induced augmentation of cocaine reward.
This study used both wild-type mice and mice with a constitutive deletion of CREB to determine whether chronic FS leads to an augmentation of cocaine CPP and whether this augmentation is dependent on CREB. Additionally, we examined the role of CRFR1 in modulating the effects of stress-induced augmentation of cocaine reward. Lastly, by determining that CRFR1 activation is required for both augmented cocaine conditioning and stress-induced phosphorylation of CREB, we demonstrate a link between CRFR1 activation and CREB as one important mechanism underlying this response.
MATERIALS AND METHODS
Animals
CREBαΔ mutant mice and wild-type littermates were bred and maintained in a F1 hybrid background (129SvEvTac;C57Bl/6). All mice (2–4 months) were group housed with food and water available ad libitum in an animal care facility on a 12-h light/dark cycle (lights on at 7:00 am) in accordance with the University of Pennsylvania Institutional Animal Care and Use Committee. Experiments were conducted between 8:00 am and 2:00 pm.
Drugs
Cocaine hydrochloride was obtained from NIDA Drug Supply (Research Triangle Park, NC). Saline (0.9%) was obtained from Sigma Aldrich, St Louis, MO. Antalarmin hydrochloride, kindly provided by Dr Kenner Rice (NIDA, NIAAA) was dissolved in warm DMSO (65°C), vortexed and diluted to a final concentration of 1.0 mg/ml in 10% DMSO in warm saline, which served as vehicle, 1 h before injections. Drugs were injected in a volume of 0.1 ml/10 g of body weight.
Behavioral Experiments
Forced swim
Mice were placed in plastic cylinders (23 cm tall × 14 cm-diameter) containing water (23–25°C), 10 cm deep for 6 min. Mice were exposed to FS once daily for 7 consecutive days. Total immobility time was scored daily by a trained observer for all experiments using CREBαΔ mutant mice (Figure 3a). When mice were injected with antalarmin or vehicle (Figure 5a), the FS score was assessed using the ViewPoint videotracking system (View point S.A., Champagne au Mont d'Or, France) and confirmed with visual scoring by a trained observer.
CPP after repeat FS exposure
(Figure 1) Preconditioning phase: On day 1, mice were allowed to explore both sides of an unbiased two chambered CPP apparatus (20 × 20 × 20 cm) for 900 s and time spent in each side was recorded. These data were used to separate animals into groups with approximately equal preference for each side and any individual bias was not considered when assigning drug or side pairings. Repeat FS exposure: Animals were exposed to 6 min FS for 7 consecutive days. In a separate series of experiments, animals were administered antalarmin (10 mg/kg, i.p.) or vehicle 30 min before each FS exposure. Conditioning phase. Beginning on day 9, after exposure to the repeated FS, animals were paired for 8 days, with the saline group receiving injections (0.9% sodium chloride) on both sides of the boxes, whereas the drug-paired group received cocaine (10 mg/kg) on one side and saline on the other side. Drug-paired sides were randomized among all groups. Testing phase: On day 17, animals were all given a saline injection and allowed to explore freely between the two sides and time spent on each side was recorded. The Preference Score (time spent in drug-paired side minus time in saline-paired side) was calculated for each mouse.

Experimental paradigm for Cocaine CPP Preconditioning day (PRE;1): On day 1, mice were tested for initial bias. Forced swim stress: mice were exposed to forced swim stress for 6 min/day; days 2–8). Thirty minutes before each forced swim exposure separate cohorts of mice were injected with antalarmin (arrows; 10 mg/kg) or saline. Conditioning phase (days 9–16): The distinct sides of conditioning boxes were paired with either saline (S) or cocaine (C; 10 mg/kg, i.p.) injections, with one exposure to box per 24 h. Test (day 17): Mice were tested for preference.
Behavioral data analysis
For FS data, the time immobile was scored as described above and analyzed using repeated measures ANOVAs with time, stress, and pretreatment (or genotype) as factors. For all CPP experiments, the preference scores (described above) were analyzed for all groups using repeated measures ANOVAs with drug, stress, and pretreatment (or genotype) as factors. Post hoc analyses for all behavioral experiments were conducted using the Fisher's PLSD post hoc test.
Phospho-CREB Immunohistochemistry
A separate cohort of mice was exposed to repeat FS. Twenty-four hours after last exposure, mice were deeply anesthetized with sodium pentobarbital (50 mg/kg, i.p.) and transcardially perfused with 30 ml of phosphate buffered saline (PBS) followed by 40 ml of 4% paraformaldehyde (PFA) in PBS. The brains were removed and placed in 4% PFA overnight at 4°C, followed by immersion in a solution of 30% sucrose in PBS containing 0.1% sodium azide at 4°C for at least 48 h. Brains were frozen and 40 μm sections were placed in PBS with 0.1% sodium azide and stored at 4°C. Before immunohistochemical labeling, sections were incubated for 20 min in 0.75% H2O2 in PBS followed by several rinses in PBS. Sections were then rinsed several times with PBS containing 0.3% Triton X-100 (PBST), 0.04% bovine serum albumin (BSA) before incubation in rabbit anti-pCREB (Abcam, Cambridge, MA). Sections were incubated for 3 days at 4°C in pCREB primary antisera (1 : 1000) diluted in PBST+BSA containing 0.1% sodium azide. Sections were rinsed several times in PBST+BSA before 90 min incubation in secondary antisera (1 : 200, biotinylated donkey anti-rabbit; Jackson ImmunoResearch, West Grove, PA). After additional rinses, sections were incubated in avidin–biotin complex (ABC elite kit, Vector Laboratories, Burlingame, CA) for 90 min. After PBS rinses, sections were incubated for 5 min in 0.04% 3,3′-diaminobenzidine-4HCl (DAB; Sigma, St Louis, MO) containing 0.01% H2O2 and 0.06% nickel sulfate in Tris Buffer for a black reaction product that was terminated by rinses in PBS. After processing, sections were mounted on glass slides, dehydrated, and coverslipped. Immunoreactivity was visualized using a Nikon Eclipse E600 microscope (Melville, NY) and images were captured with a QImaging Retiga 1300 (Surrey, British Columbia, Canada) using Image-Pro Plus software (MediaCybernetics, Bethesda, MD).
Immunohistochemical data analysis
pCREB-immunolabeled profiles from each brain region were quantified bilaterally from at least two sections per mouse (leading to four counts per animal) and averaged. The person quantifying was blind to group assignments. Anatomical regions were identified according to the stereotaxic atlas of Franklin and Paxinos (Franklin and Paxinos, 2007). Single-labeled images were quantified using ImageJ software. The number of single-labeled cells in each brain region was determined for each animal. Group differences in pCREB immunoreactivity were assessed using two-way ANOVAs with drug (antalarmin vs saline) and stress (no stress vs FS) as the independent variables and number of cells as the dependent variable. Fisher's post hoc comparisons were conducted when main effects or interactions were present.
Plasma corticosterone measurement
Trunk blood was collected in heparinized tubes from separate groups of animals 20 min after FS exposure and at an equivalent time for non-stress (NS) controls. Plasma was separated by centrifugation and stored at –20°C until assayed. Plasma corticosterone was measured by radioimmunoassay using a commercially available kit (MP Biochemicals Inc., Irvine, CA). Intra-assay coefficient of variation was below 20%.
Data analysis for corticosterone levels
The data were analyzed using individual two-way ANOVAs for each day, with stress and either pretreatment or genotype as the main factors and Fisher's PLSD test for post hoc analyses.
RESULTS
Chronic FS-Augmentation of Cocaine Place Preference is Dependent on CREB
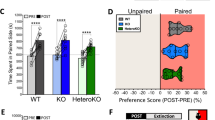
Repeated exposure to FS before cocaine conditioning augmented place preference in wild-type mice. As shown in Figure 2, the wild-type mice that were previously exposed to repeated FS showed a significantly greater preference for the side paired with cocaine ((F(1,39)=45.485), p<0.0001; effect of drug; Fisher's post hoc test, p<0.05; WT/FS/COC vs WT/NS/COC). Although all cocaine groups exhibited a significant preference compared with their respective saline controls (Figure 2; effect of drug, p<0.0001; Fisher's post hoc p<0.05 for all cocaine groups compared with respective saline control), this augmentation after FS exposure was not seen in the CREBαΔ mutant mice. In addition, we found that at a low dose of cocaine (2.5 mg/kg), neither WT nor CREB mutant mice showed CPP, regardless of stress history (Supplementary Figure S1). A higher dose of cocaine (5 mg/kg) did not elicit CPP in non-stressed (WT/NS) mice; however, it did elicit significant place preference in the WT animals that were previously stressed (WT/FS), showing a potentiation of cocaine reward (effect of drug, interaction; Fisher's post hoc test p<0.05; WT/NS/5.0 vs WT/FS/5.0). CREB mutant mice showed a significant place preference for cocaine at 5 mg/kg (MUT/NS) compared with WT animals, as has been demonstrated earlier (Fisher's post hoc test p<0.05) (Walters and Blendy, 2001). However, previous exposure to stress (MUT/FS) did not lead to increased place preference to cocaine. For all subsequent experiments, we used the dose of cocaine (10 mg/kg), which elicited a significant place preference to cocaine that was similar in magnitude for both genotypes, thereby conferring an equivalent baseline with which to compare subsequent stressor and drug effects.

Wild-type but not CREB mutant mice show augmented conditioned cocaine place preference after chronic FS. Both wild-type and mutant mice paired with cocaine (WT/NS, MUT/NS) showed a significant place preference to the cocaine-paired side on test day. Wild-type mice exposed to forced swim (WT/FS) showed increased preference scores (time on paired side minus time on unpaired side); CREB mutant mice did not show this stress-induced potentiation (MUT/FS). Data are expressed as mean±SEM; six mice per group. *p<0.05 from corresponding saline group; **p<0.05 from cocaine non-stress group (ANOVA; Fisher's post hoc test)
Despite the differences in immobility on days 1, 2 and 5 (Figure 3a; p<0.05 post hoc analyses WT vs MT), repeated FS exposure led to a general increase in immobility over time in both genotypes (Figure 3a; effect of time (F1,90=20.99); p<0.0001). Specifically, both wild-type and mutant mice showed an equivalent elevated amount of immobility on the 2 days before cocaine conditioning. Both wild-type and mutant animals showed similar baseline plasma corticosterone levels and similar increases in corticosterone after FS exposure (Figure 3b; effect of stress, day 1: (F1,25=34.998); p<0.0001 and day 7: (F1,18=6.772), p<0.01). Therefore, although initially, there might be a difference in stress reactivity between the two groups, there is no change in behavior or coping strategy several days leading up to cocaine conditioning.

Wild-type and CREB mutant mice show equivalent forced swim behavior and corticosterone response to FS. (a) CREB mutant mice (MUT) showed decreased immobility on days 1, 2 and 5 of the forced swim exposure. However, immobility increased over the 7 days of FS testing in both wild-type (WT) and CREB mutant mice. (b) Wild-type and CREB mutant mice exhibited similar changes in plasma corticosterone after acute as well as repeated FS stress exposures. Additionally, both groups showed similar basal levels of corticosterone in the non-stressed groups. Data are expressed as mean±SEM; 6–9 mice per group. *p<0.05 from either corresponding mutant group (a) or corresponding non-stress group (b) (ANOVA; Fisher's post hoc test).
Chronic FS-Augmentation of Cocaine Place Preference is Dependent on CRFR1 Activation
In preliminary experiments, we found that treatment with lower doses of the CRFR1 antagonist antalarmin (5 and 7.5 mg/kg) significantly increased plasma corticosterone levels in non-stressed mice (725±34 ng/ml and 585±33 ng/ml) when compared with vehicle injection (176±64 ng/ml), whereas a dose of 10 mg/kg did not significantly increase basal corticosterone levels (334±44 ng/ml). Therefore, we chose this dose to determine whether antalarmin modulates stress-induced cocaine reward. As shown earlier, saline pretreated mice exposed to repeated FS showed a significantly greater preference for the side paired with cocaine (Figure 4; (F1,40=45.368) effect of drug, p<0.05 for all ANOVAs; Fisher's post hoc test p<0.05 SAL/NS/COC vs SAL/FS/COC). In contrast, mice pretreated with antalarmin did not show the FS-induced increase in cocaine CPP (Fisher's post hoc test p>0.05; ANT/NS/COC vs ANT/FS/COC; p<0.001 ANT/FS/COC vs SAL/FS/COC). Additionally, neither stress nor antalarmin pretreatment altered place preference scores in saline conditioned mice, which were close to zero for all groups.

Antalarmin pretreatment during repeat FS exposure blocks stress-induced potentiation of cocaine place preference. Wild-type mice were conditioned to cocaine (10 mg/kg). All mice conditioned to cocaine showed a place preference regardless of pretreatment. Mice paired with cocaine and previously exposed to the forced swim showed an augmentation of place preference. Mice pretreated with antalarmin (10 mg/kg) showed similar cocaine place preference regardless of previous FS exposure. Data are expressed as mean±SEM; 6 mice per group. *p<0.05 from corresponding saline group; **p<0.05 from cocaine non-stress group (ANOVA; Fisher's PLSD post hoc test).
Just as in the CREBαΔ mice, the ability of antalarmin to block FS-induced augmentation of cocaine reward may not be dependent on changes in immobility behavior in the FS (Figure 5a). Repeated FS exposure led to an increase in time spent immobile in both pretreatment groups ((F1,60=7.53); p<0.0001). Antalarmin pretreatment (10 mg/kg) blocked the ability of stress to increase plasma corticosterone levels on day 1 (Figure 5b; interaction (F1,16=8.932); p<0.01, Fisher's post hoc test p<0.05; ANT/FS vs SAL/FS). However, after repeated FS exposure, both saline- and antalarmin-treated mice exhibited comparable increases in plasma corticosterone in response to stress (Figure 5b; effect of stress (F 1,17=14.759), p<0.01).

Antalarmin- and vehicle-treated mice show equivalent forced swim behavior and corticosterone response to FS. (a) Wild-type mice given saline injections (VEH) showed increased immobility over 7 days of repeated forced swim. Pretreatment with antalarmin (ANT) injection did not alter immobility behavior of wild-type mice at any time point. Both groups showed a significant increase in immobility over 7 days. (b) Forced swim exposure increased corticosterone levels both acutely as well as after repeated exposure. Wild-type mice injected with saline (VEH) showed increased corticosterone levels after acute FS exposure. Pretreatment with antalarmin blocked the forced swim stress-induced increase in plasma corticosterone in wild-type mice (VEH/FS; ANT/FS) after the first, but not after repeated exposures. In addition, antalarmin injection does not alter basal corticosterone levels in the non-stressed groups (NS). Data are expressed as mean±SEM; 4–6 mice per group. *p<0.05 from corresponding non-stress group (ANOVA; Fisher's PLSD post hoc test).
Stress-Elicited pCREB Expression in Forebrain Regions After Chronic FS is Dependent on CRFR1 Receptor Activation
To further elucidate the downstream signaling involved in FS-induced augmentation of cocaine reward, and to begin to link the effects of CRFR1 receptor activation with CREB, we examined CREB phosphorylation in a variety of brain regions after chronic FS. Twenty-four hours after repeated exposure to FS (the time point at which mice are first exposed to cocaine CPP), pCREB-immunoreactivity is increased in the lateral septum (LS), nucleus accumbens (NAc) core and shell, ventral and dorsal bed nucleus of the stria terminalis (vBNST, dBNST), and basolateral and central nuclei of the amygdala (BLA, CeA; Figure 6). However, in mice pretreated with antalarmin, this stress-induced increase in pCREB-immunoreactivity is blunted in the LS and the NAc core and shell (Figure 6b-d; effect of drug, stress, and interaction, p<0.05 for all ANOVAs, Fisher's post hoc tests: saline FST vs antalarmin FST, p<0.05). Not all areas that show increased pCREB after FS were affected by antalarmin pretreatment, as comparable increases were seen in the vBNST, dBNST, BLA, and CeA in antalarmin and saline pretreated mice (Figure 6e-h; effect of stress, p<0.001 for all ANOVAs). Antalarmin did not alter basal pCREB-immunoreactivity in NS controls in any brain regions except for the NAc core.

Stress-elicited pCREB expression in certain forebrain regions is dependent on CRF1 receptor activation. (a) Representative micrographs show pCREB expression in lateral septum and nucleus accumbens core (NAc core) after chronic FS exposure (FST Sal) and antalarmin treatment (FST Ant). Saline injected mice exhibited an increase in pCREB-immunoreactive profiles after repeated stress exposure in lateral septum (LS) (b), the nucleus accumbens (NAc) core (c), and shell (d), ventral and dorsal bed nucleus of the stria terminalis (vBNST, dBNST) (e, f), and basolateral and central nuclei of the amygdala (BLA, CeA) (g, h). Mice pretreated with antalarmin did not exhibit stress-induced increases in pCREB protein expression in the LS, NAc core and NAc shell (b-d). Stress-induced increases in pCREB activation remained intact in vBNST, dBNST, BLA and CeA (e-h) after antalarmin pretreatment. Data are expressed as mean±SEM. **p<0.01, ***p<0.001 from non stress group; #p<0.05, ###p<0.001 from saline treated group (ANOVA; Bonferroni–Dunn post hoc test).
DISCUSSION
Drug addiction is a disease hallmarked by cyclical changes in drug taking behavior, with initial drug use often leading to chronic abuse and abstinence from drug use frequently leading to relapse. Exposure to stress can alter vulnerability to drug abuse at each of these stages. But exactly how stress alters the rewarding properties of cocaine and what components of the stress response are involved is unknown. Previous work has demonstrated a role for the transcription factor CREB in mediating interactions between stress and reactivation of cocaine conditioning. In addition, modulating CRFR1 activity can alter drug taking and reward as well as the interaction between stress and addiction. Therefore, these molecules are appropriate targets to investigate as possible mediators for stress potentiation of drug reward.
Augmentation of Cocaine Reward After Chronic FS
The present data demonstrate that repeated FS exposure over 7 days, 24 h before the first cocaine conditioning day potentiates cocaine CPP. This model may have important clinical implications, as chronic stress is thought to be a critical factor underlying the vulnerability to drug abuse. Our data coincides with studies that show that other stressors such as shock, tail pinch, or social aggression can enhance the reinforcing properties of cocaine as assessed by acquisition of cocaine self administration (Goeders and Guerin, 1994; Haney et al, 1995; Miczek and Mutschler, 1996; Piazza et al, 1990; Shaham et al, 1995; Shaham et al, 1994a). However, very few studies have examined the effects of stress in the development of cocaine CPP when stress precedes any exposure to drug. An earlier study has shown that FS exposure during the conditioning phase of the experiment augments cocaine place preference (McLaughlin et al, 2003). In that study, mice exposed to FS are administered cocaine at a time when corticosterone levels are likely elevated above baseline (Conti et al, 2002). In contrast, in the present studies, cocaine conditioning takes place when corticosterone levels are back to NS, baseline values (24 h after the last FS exposure) suggesting factors other than increased corticosterone may underlie the augmented cocaine place preference observed in these studies. Indeed, the role of corticosterone in mediating CPP remains equivocal. Although Li et al (2007) have recently shown that corticosterone or chronic footshock can potentiate morphine CPP, Dietz et al (2007) have demonstrated that corticosterone itself is neither rewarding nor aversive. In addition, Der-Avakian et al (2005, 2007) showed that blocking corticosterone during a stressor presentation (ie before conditioning) has no effect on potentiation of subsequent morphine reward; however, corticosterone inhibition during conditioning blocks this potentiation. Together, these studies demonstrate that changes in corticosterone, before conditioning do not affect subsequent stress-induced potentiation of conditioned reward, suggesting that the effect of stress may be mediated by other factors. Moreover, these factors/effects appear to be long lasting as manifestation of this stress on conditioned reward is evident well after cessation of stress exposure.
Role of CRFR1 Signaling in Chronic FS-Induced Augmentation of Cocaine Reward
Similar to activation of the HPA axis, the extra-hypothalamic CRF system is also activated after exposure to stressors. Alterations in this system, specifically by the CRFR1 receptor, have been shown to be critical for stress-induced changes in cocaine reward (Goeders, 1997; Goeders et al, 1990). However, the role of the CRF system in mediating the development of cocaine conditioning after chronic stress is largely unknown. To determine whether CRFR1 signaling is important in mediating FS-induced potentiation of cocaine reward, we pretreated wild-type animals with antalarmin, a specific non-peptide CRFR1 antagonist (Webster et al, 1996), and assessed alterations in reward. Administration of antalarmin before each FS exposure blocks FS-induced potentiation of cocaine place preference in wild-type animals, demonstrating for the first time that the CRFR1 mediates stress-induced potentiation of subsequent cocaine reward.
Several studies have shown a role for CRFR1 in conditioning to cocaine and morphine. Intracerebroventricular administration of a CRFR1 antagonist blocks cocaine locomotion and acquisition of cocaine CPP when given before each conditioning trial (Lu et al, 2003). In addition, CRFR1 antagonist treatment decreases maintenance of cocaine self-administration (Goeders and Guerin, 2000). These studies suggest that CRFR1 activation is critical during the conditioning procedure. However, in our studies, administration of antalarmin occurs before conditioning procedure and does not affect conditioning to cocaine. Therefore, blocking CRFR1 activation during stressor exposure before conditioning identifies a specific role for this receptor only during a stressful experience.
FS-Induced Augmentation of Cocaine Reward is not Dependent on Behavioral or Endocrine Responses
Altered behavioral or endocrine responses to FS stress in CREB-deficient mice or in wild-type mice pretreated with antalarmin may underlie the changes in stress-induced reward behaviors observed in these mice. Therefore, we quantified the immobility time in the FS over repeated exposures and corticosterone responses to this stress. Immobility behavior in the FS may be a form of adaptation from an inescapable stress, which is interrupted with bouts of activity and escape motivated behavior. As seen earlier, CREB mutant animals show an initial decrease in immobility when compared with the wild-type mice (Conti et al, 2002). However, this alteration in immobility is not seen after antalarmin treatment. This difference in behavioral immobility may be due to several reasons. First, it is possible that CRFR1 receptor coupling to CREB is not uniform throughout the brain. Thus, despite the effect we see on stress-induced augmentation that connects CREB and CRFR1, other behavioral outcome measures, such as immobility in the FS, may not be mediated by this interaction. Alternatively, this altered baseline response in CREB mutant animals may be due to developmental compensations for the constitutive loss of CREB and may not necessarily be related to the role of CREB as a putative downstream signaling molecule for CRF. Finally, antalarmin has also been shown to have mixed effects on immobility in the FS test, with data demonstrating decreased immobility (Griebel et al, 2002; Land et al, 2008) as well as no change in behavior (Jutkiewicz et al, 2005). Thus, immobility differences may be more apparent in different strains or species.
Importantly, independent of initial immobility behavior, by day 7, all groups showed an increased immobility such that there was no difference between wild-type and CREB mutant animals or saline-treated and antalarmin-treated animals. Thus, immediately before conditioning, the behavioral responsivity of all groups of mice remained equivalent to their control counterparts, implying that immobility behavior in the FS does not contribute substantially to stress-induced potentiation of cocaine conditioning in our model.
Behavioral reactivity to stress may be distinct from physiological changes due to stressor exposure and these may also affect subsequent behavior. Although FS exposure increased corticosterone release in mice, only an acute administration of antalarmin blocked corticosterone increase after FS. This effect was not observed after repeated FS exposure. In addition, there were no differences in corticosterone release after single or repeated FS exposure in CREB mutant mice when compared with wild-type mice. Together with earlier studies discussed earlier, these data indicate that corticosterone changes during stressor exposure in advance of conditioning do not contribute to stress-induced potentiation of cocaine reward.
Importance of CREB in the Ability of Chronic FS to Potentiate Cocaine CPP
The current studies demonstrate that CREB function is necessary for chronic stress to augment cocaine reward. This CREB requirement for stress–drug interaction, closely parallels what has been demonstrated earlier in stress-induced reinstatement in these mice (Kreibich and Blendy, 2004). As CREB is constitutively deleted in these mice, the locus where CREB is acting to facilitate stress-induced augmentation of cocaine reward is unknown. Wild-type mice exhibit increase in pCREB levels in many brain regions after chronic FS. Several of these regions have been implicated in both addiction and stress circuitry, including the core and shell of the NAc, the LS, BNST, and the amygdala. Thus, CREB may be critical in a number of brain regions or perhaps within the entire circuit mediating these stress–drug interactions.
CRFR1 Activation is Upstream of CREB Signaling in FS-Induced Potentiation of Cocaine Reward Circuitry
The CRFR1 is coupled to the stimulatory G protein Gαs and can thus activate PKA and subsequently CREB (Kasagi et al, 2002). We found that treatment with a CRFR1 antagonist before stress exposure blocked the ability of FS to lead to increased levels of pCREB within a discrete set of brain regions. Repeated exposure to the FS-induced elevated pCREB levels in several areas of the brain, including the NAcNAc, LS, BNST, and amygdala. All of these extrahypothalamic brain regions show neural activation after acute FS exposure (Dayas et al, 2001; Duncan et al, 2009; Duncan et al, 1993; Kreibich and Blendy, 2004). However, the specific role of CRFR1 in mediating responses to FST in each of these regions is unknown. Although several brain nuclei showed increased levels of pCREB expression after chronic FS, pretreatment with antalarmin reduced this pCREB expression only in the LS and the NAc. As antalarmin pretreatment also blocked the ability of repeated stress to potentiate cocaine CPP, this suggests a specific role for these nuclei in mediating the interaction between stress and cocaine reward.
The NAc and LS are interconnected nuclei that are part of the limbic circuitry influencing stress responsivity as well as motivational aspects of behavior and responses to cocaine (Carlezon and Thomas, 2009; Faure et al, 2008; Louilot et al, 1989; Sheehan et al, 2004). The NAc has an important function in mediating acquisition of incentive and appetitive learning through a cAMP-dependent pathway as well as in modulating responses to emotionally relevant stimuli (Cooper and Knutson, 2008; Kelley, 2004; Kheirbek et al, 2008; Reynolds and Berridge, 2002). In addition, CREB activity in the NAc modulates emotional behavior including anxiogenic and nociceptive responses, immobility behavior in the FS, as well as cocaine reward (Barrot et al, 2002; Carlezon et al, 1998; Pliakas et al, 2001). Specifically, disruption of CREB in the NAc by overexpression of the dominant negative form of CREB increases cocaine reward (Carlezon et al, 1998; Dinieri et al, 2009; Pliakas et al, 2001). Therefore, blocking CRFR1 before each stressor exposure, which downregulates stress-induced CREB signaling in the NAc, may allow for subsequent changes in threshold of cocaine reward, resulting in an augmented conditioned response.
Activation of the LS and changes in neurotransmitter release have also been correlated with behavioral responses to stressors, and in particular, immobility behavior in the FS (Duncan et al, 1993; Ebner et al, 1999; Kirby and Lucki, 1997). Of interest, cells in the LS are also activated after exposure to cocaine or a cocaine-paired environment (Brown et al, 1992; Franklin and Druhan, 2000; Trinh et al, 2003). Furthermore, other components of the extrahypothalamic-CRF system, such as the CRFR2 have been shown to be altered in the LS after chronic cocaine administration and withdrawal (Liu et al, 2005). Therefore LS, like the NAc is part of the neural circuitry, which mediates responses to both stress and cocaine. Both of these nuclei are activated by repeated FS exposure and this increase in pCREB levels may either lower the threshold for subsequent cocaine reward or increase motivational salience for conditioning, potentiating subsequent cocaine CPP. However, blocking CRFR1 receptors with antalarmin inhibits the activation of NAc and LS during the repeated FS exposures, thereby preventing the facilitation of stress-induced augmented conditioning. Blocking pCREB levels in these nuclei does not correlate with altered immobility behavior or corticosterone levels in the FS, which is somewhat contrary to earlier data that implicate these regions in behavioral responses to FS (Carlezon et al, 1998; Duncan et al, 1993; Ebner et al, 1999; Kirby and Lucki, 1997; Pliakas et al, 2001). However, earlier studies have not examined the sufficiency of these nuclei in mediating behavioral responses over chronic stress exposure. Thus, other brain regions, such as the hypothalamus, amygdala, or the BNST may mediate behavioral and endocrine responses to chronic FS exposure.
Although other brain regions not described here may also be involved in the stress-induced potentiation of cocaine reward, our data and the relationship between CRFR1 and pCREB point to specific roles for the NAc and the LS in mediating stress-induced potentiation of CPP. Future studies using viral vectors to block or enhance CREB in these specific brain regions will reveal whether these areas are sufficient for stress-induced potentiation of cocaine reward.
Clinical research indicates that life stress is a risk factor in the development of addiction and the severity of this stress is correlated with drug use, with higher levels of stress yielding higher levels of use (McFall et al, 1992). The current study not only provides us with a model to study the ability of stress to promote drug use but also provides insight into the molecular mechanisms underlying this phenomenon. We have demonstrated a role for CRFR1 and CREB signaling in stress-induced augmentation of cocaine reward and characterized brain regions that may identify the locus or circuitry involved in this complex interaction between stress and drug addiction. The combination of detailed molecular and system wide approaches in the future will enhance the ability to identify common receptor signaling and molecular mechanisms underlying behavioral effects that may affect both initial drug taking as well as relapse to addiction.
DISCLOSURE
The authors declare that, except for income received from primary employers, no financial support or compensation has been received from any individual or corporate entity over the past 3 years for research or professional service and there are no personal financial holdings that could be perceived as constituting a potential conflict of interest.
References
Barrot M, Olivier JD, Perrotti LI, DiLeone RJ, Berton O, Eisch AJ et al (2002). CREB activity in the nucleus accumbens shell controls gating of behavioral responses to emotional stimuli. Proc Natl Acad Sci USA 99: 11435–11440.
Brown EE, Robertson GS, Fibiger HC (1992). Evidence for conditional neuronal activation following exposure to a cocaine-paired environment: role of forebrain limbic structures. J Neurosci 12: 4112–4121.
Carlezon Jr WA, Thomas MJ (2009). Biological substrates of reward and aversion: a nucleus accumbens activity hypothesis. Neuropharmacology 56 (Suppl 1): 122–132.
Carlezon Jr WA, Thome J, Olson VG, Lane-Ladd SB, Brodkin ES, Hiroi N et al (1998). Regulation of cocaine reward by CREB. Science (New York, NY) 282: 2272–2275.
Conti AC, Cryan JF, Dalvi A, Lucki I, Blendy JA (2002). cAMP response element-binding protein is essential for the upregulation of brain-derived neurotrophic factor transcription, but not the behavioral or endocrine responses to antidepressant drugs. J Neurosci 22: 3262–3268.
Cooper JC, Knutson B (2008). Valence and salience contribute to nucleus accumbens activation. NeuroImage 39: 538–547.
Dayas CV, Buller KM, Crane JW, Xu Y, Day TA (2001). Stressor categorization: acute physical and psychological stressors elicit distinctive recruitment patterns in the amygdala and in medullary noradrenergic cell groups. Eur J Neurosci 14: 1143–1152.
Der-Avakian A, Bland ST, Rozeske RR, Tamblyn JP, Hutchinson MR, Watkins LR et al (2007). The effects of a single exposure to uncontrollable stress on the subsequent conditioned place preference responses to oxycodone, cocaine, and ethanol in rats. Psychopharmacology (Berl) 191: 909–917.
Der-Avakian A, Will MJ, Bland ST, Deak T, Nguyen KT, Schmid MJ et al (2005). Surgical and pharmacological suppression of glucocorticoids prevents the enhancement of morphine conditioned place preference by uncontrollable stress in rats. Psychopharmacology (Berl) 179: 409–417.
Dietz D, Wang H, Kabbaj M (2007). Corticosterone fails to produce conditioned place preference or conditioned place aversion in rats. Behav Brain Res 181: 287–291.
Dinieri JA, Nemeth CL, Parsegian A, Carle T, Gurevich VV, Gurevich E et al (2009). Altered sensitivity to rewarding and aversive drugs in mice with inducible disruption of cAMP response element-binding protein function within the nucleus accumbens. J Neurosci 29: 1855–1859.
Duncan GE, Inada K, Farrington JS, Koller BH, Moy SS (2009). Neural activation deficits in a mouse genetic model of NMDA receptor hypofunction in tests of social aggression and swim stress. Brain Res 1265: 186–195.
Duncan GE, Johnson KB, Breese GR (1993). Topographic patterns of brain activity in response to swim stress: assessment by 2-deoxyglucose uptake and expression of Fos-like immunoreactivity. J Neurosci 13: 3932–3943.
Ebner K, Wotjak CT, Holsboer F, Landgraf R, Engelmann M (1999). Vasopressin released within the septal brain area during swim stress modulates the behavioural stress response in rats. Eur J Neurosci 11: 997–1002.
Faure A, Reynolds SM, Richard JM, Berridge KC (2008). Mesolimbic dopamine in desire and dread: enabling motivation to be generated by localized glutamate disruptions in nucleus accumbens. J Neurosci 28: 7184–7192.
Franklin KBJ, Paxinos G (2007). The Mouse Brain in Stereotaxic Coordinates. 3rd edn. Academic Press: San Diego.
Franklin TR, Druhan JP (2000). Expression of Fos-related antigens in the nucleus accumbens and associated regions following exposure to a cocaine-paired environment. Eur J Neurosci 12: 2097–2106.
Goeders NE (1997). A neuroendocrine role in cocaine reinforcement. Psychoneuroendocrinology 22: 237–259.
Goeders NE, Bienvenu OJ, De Souza EB (1990). Chronic cocaine administration alters corticotropin-releasing factor receptors in the rat brain. Brain Res 531: 322–328.
Goeders NE, Guerin GF (1994). Non-contingent electric footshock facilitates the acquisition of intravenous cocaine self-administration in rats. Psychopharmacology (Berl) 114: 63–70.
Goeders NE, Guerin GF (2000). Effects of the CRH receptor antagonist CP-154, 526 on intravenous cocaine self-administration in rats. Neuropsychopharmacology 23: 577–586.
Griebel G, Simiand J, Steinberg R, Jung M, Gully D, Roger P et al (2002). 4-(2-Chloro-4-methoxy-5-methylphenyl)-N-[(1S)-2-cyclopropyl-1-(3-fluoro-4-methylphenyl)ethyl]5-methyl-N-(2-propynyl)-1, 3-thiazol-2-amine hydrochloride (SSR125543A), a potent and selective corticotrophin-releasing factor(1) receptor antagonist. II. Characterization in rodent models of stress-related disorders. J Pharmacol Exp Ther 301: 333–345.
Hadaway PF, Alexander BK, Coambs RB, Beyerstein B (1979). The effect of housing and gender on preference for morphine-sucrose solutions in rats. Psychopharmacology (Berl) 66: 87–91.
Haney M, Maccari S, Le Moal M, Simon H, Piazza PV (1995). Social stress increases the acquisition of cocaine self-administration in male and female rats. Brain Res 698: 46–52.
Jutkiewicz EM, Wood SK, Houshyar H, Hsin LW, Rice KC, Woods JH (2005). The effects of CRF antagonists, antalarmin, CP154 526, LWH234, and R121919, in the forced swim test and on swim-induced increases in adrenocorticotropin in rats. Psychopharmacology 180: 215–223.
Kaplan HB, Martin SS, Johnson RJ, Robbins CA (1986). Escalation of marijuana use: application of a general theory of deviant behavior. J Health Soc Behav 27: 44–61.
Kasagi Y, Horiba N, Sakai K, Fukuda Y, Suda T (2002). Involvement of cAMP-response element binding protein in corticotropin-releasing factor (CRF)-induced down-regulation of CRF receptor 1 gene expression in rat anterior pituitary cells. J Neuroendocrinol 14: 587–592.
Kelley AE (2004). Ventral striatal control of appetitive motivation: role in ingestive behavior and reward-related learning. Neurosci Biobehav Rev 27: 765–776.
Kheirbek MA, Beeler JA, Ishikawa Y, Zhuang X (2008). A cAMP pathway underlying reward prediction in associative learning. J Neurosci 28: 11401–11408.
Kirby LG, Lucki I (1997). Interaction between the forced swimming test and fluoxetine treatment on extracellular 5-hydroxytryptamine and 5-hydroxyindoleacetic acid in the rat. J Pharmacol Exp Ther 282: 967–976.
Koob G, Kreek MJ (2007). Stress, dysregulation of drug reward pathways, and the transition to drug dependence. Am J Psychiatry 164: 1149–1159.
Kreibich AS, Blendy JA (2004). cAMP response element-binding protein is required for stress but not cocaine-induced reinstatement. J Neurosci 24: 6686–6692.
Kwon MS, Seo YJ, Shim EJ, Choi SS, Lee JY, Suh HW (2006). The effect of single or repeated restraint stress on several signal molecules in paraventricular nucleus, arcuate nucleus and locus coeruleus. Neuroscience 142: 1281–1292.
Land BB, Bruchas MR, Lemos JC, Xu M, Melief EJ, Chavkin C (2008). The dysphoric component of stress is encoded by activation of the dynorphin kappa-opioid system. J Neurosci 28: 407–414.
Li Y, Li GY, Li LJ, Wang CH, Li ZX, Zhang JL et al (2007). Subsequently enhanced CPP to morphine following chronic but not acute footshock stress associated with corticosterone mechanism in rats. Int J Neurosci 117: 1237–1255.
Liu J, Yu B, Orozco-Cabal L, Grigoriadis DE, Rivier J, Vale WW et al (2005). Chronic cocaine administration switches corticotropin-releasing factor2 receptor-mediated depression to facilitation of glutamatergic transmission in the lateral septum. J Neurosci 25: 577–583.
Louilot A, Taghzouti K, Simon H, Le Moal M (1989). Limbic system, basal ganglia, and dopaminergic neurons. Executive and regulatory neurons and their role in the organization of behavior. Brain Behav Evol 33: 157–161.
Lu L, Liu Z, Huang M, Zhang Z (2003). Dopamine-dependent responses to cocaine depend on corticotropin-releasing factor receptor subtypes. J Neurochem 84: 1378–1386.
Maddahian E, Newcomb MD, Bentler PM (1988a). Adolescent drug use and intention to use drugs: concurrent and longitudinal analyses of four ethnic groups. Addict Behav 13: 191–195.
Maddahian E, Newcomb MD, Bentler PM (1988b). Risk factors for substance use: ethnic differences among adolescents. J Subst Abuse 1: 11–23.
McEvoy AN, Bresnihan B, Fitzgerald O, Murphy EP (2002). Corticotropin-releasing hormone signaling in synovial tissue vascular endothelium is mediated through the cAMP/CREB pathway. Ann N Y Acad Sci 966: 119–130.
McFall ME, Mackay PW, Donovan DM (1992). Combat-related posttraumatic stress disorder and severity of substance abuse in Vietnam veterans. J Stud Alcohol 53: 357–363.
McLaughlin JP, Marton-Popovici M, Chavkin C (2003). Kappa opioid receptor antagonism and prodynorphin gene disruption block stress-induced behavioral responses. J Neurosci 23: 5674–5683.
Miczek KA, Mutschler NH (1996). Activational effects of social stress on IV cocaine self-administration in rats. Psychopharmacology (Berl) 128: 256–264.
Piazza PV, Deminiere JM, le Moal M, Simon H (1990). Stress- and pharmacologically-induced behavioral sensitization increases vulnerability to acquisition of amphetamine self-administration. Brain Res 514: 22–26.
Piazza PV, Le Moal M (1998). The role of stress in drug self-administration. Trends Pharmacol Sci 19: 67–74.
Pliakas AM, Carlson RR, Neve RL, Konradi C, Nestler EJ, Carlezon Jr WA (2001). Altered responsiveness to cocaine and increased immobility in the forced swim test associated with elevated cAMP response element-binding protein expression in nucleus accumbens. J Neurosci 21: 7397–7403.
Ramsey NF, Van Ree JM (1993). Emotional but not physical stress enhances intravenous cocaine self-administration in drug-naive rats. Brain Res 608: 216–222.
Reynolds SM, Berridge KC (2002). Positive and negative motivation in nucleus accumbens shell: bivalent rostrocaudal gradients for GABA-elicited eating, taste ‘liking’/‘disliking’ reactions, place preference/avoidance, and fear. J Neurosci 22: 7308–7320.
Sarnyai Z, Shaham Y, Heinrichs SC (2001). The role of corticotropin-releasing factor in drug addiction. Pharmacol Rev 53: 209–243.
Shaham Y, Kelsey JE, Stewart J (1995). Temporal factors in the effect of restraint stress on morphine-induced behavioral sensitization in the rat. Psychopharmacology (Berl) 117: 102–109.
Shaham Y, Rodaros D, Stewart J (1994a). Reinstatement of heroin-reinforced behavior following long-term extinction: implications for the treatment of relapse to drug taking. Behav Pharmacol 5: 360–364.
Shaham Y, Stewart J (1994b). Exposure to mild stress enhances the reinforcing efficacy of intravenous heroin self-administration in rats. Psychopharmacology (Berl) 114: 523–527.
Sheehan TP, Chambers RA, Russell DS (2004). Regulation of affect by the lateral septum: implications for neuropsychiatry. Brain Res Brain Res Rev 46: 71–117.
Sinha R (2007). The role of stress in addiction relapse. Curr Psychiatry Rep 9: 388–395.
Trinh JV, Nehrenberg DL, Jacobsen JP, Caron MG, Wetsel WC (2003). Differential psychostimulant-induced activation of neural circuits in dopamine transporter knockout and wild type mice. Neuroscience 118: 297–310.
Valverde O, Mantamadiotis T, Torrecilla M, Ugedo L, Pineda J, Bleckmann S et al (2004). Modulation of anxiety-like behavior and morphine dependence in CREB-deficient mice. Neuropsychopharmacology 29: 1122–1133.
Walters CL, Blendy JA (2001). Different requirements for cAMP response element binding protein in positive and negative reinforcing properties of drugs of abuse. J Neurosci 21: 9438–9444.
Walters CL, Cleck JN, Kuo YC, Blendy JA (2005). Mu-opioid receptor and CREB activation are required for nicotine reward. Neuron 46: 933–943.
Webster EL, Lewis DB, Torpy DJ, Zachman EK, Rice KC, Chrousos GP (1996). In vivo and in vitro characterization of antalarmin, a nonpeptide corticotropin-releasing hormone (CRH) receptor antagonist: suppression of pituitary ACTH release and peripheral inflammation. Endocrinology 137: 5747–5750.
Acknowledgements
This work was supported by the National Institute on Drug Abuse NIDA Grant DA116–49–01A2 (J.A.B.). A portion of this work was supported by the NIH Intramural Research Program of the National Institute of Diabetes and Digestive and Kidney Diseases, the National Institute on Drug Abuse and the National Institute on Alcohol Abuse and Alcoholism.
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website (http://www.nature.com/npp)
Supplementary information
Rights and permissions
About this article
Cite this article
Kreibich, A., Briand, L., Cleck, J. et al. Stress-Induced Potentiation of Cocaine Reward: A Role for CRFR1 and CREB. Neuropsychopharmacol 34, 2609–2617 (2009). https://doi.org/10.1038/npp.2009.91
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2009.91
Keywords
This article is cited by
-
Vulnerability factors for mephedrone-induced conditioned place preference in rats—the impact of sex differences, social-conditioning and stress
Psychopharmacology (2021)
-
Enduring effects of adolescent ketamine exposure on cocaine- and sucrose-induced reward in male and female C57BL/6 mice
Neuropsychopharmacology (2020)
-
Riluzole Attenuates L-DOPA-Induced Abnormal Involuntary Movements Through Decreasing CREB1 Activity: Insights from a Rat Model
Molecular Neurobiology (2019)
-
Social rank-associated stress vulnerability predisposes individuals to cocaine attraction
Scientific Reports (2018)
-
Behavioral and Biochemical Impact of Chronic Unpredictable Mild Stress on the Acquisition of Nicotine Conditioned Place Preference in Rats
Molecular Neurobiology (2018)
