





Abstract
Plant stress studies are more and more based on gene expression. The analysis of gene expression requires sensitive, precise, and reproducible measurements for specific mRNA sequences. Real-time RT-PCR is at present the most sensitive method for the detection of low abundance mRNA. To avoid bias, real-time RT-PCR is referred to one or several internal control genes, which should not fluctuate during treatments. Here, the non-regulation of seven housekeeping genes (β-tubulin, cyclophilin, actin, elongation factor 1-α (ef1α), 18S rRNA, adenine phosphoribosyl transferase (aprt), and cytoplasmic ribosomal protein L2) during biotic (late blight) and abiotic stresses (cold and salt stress) was tested on potato plants using geNorm software. Results from the three experimental conditions indicated that ef1α was the most stable among the seven tested. The expression of the other housekeeping genes tested varied upon stress. In parallel, a study of the variability of expression of hsp20.2, shown to be implicated in late blight stress, was realized. The relative quantification of the hsp20.2 gene varied according to the internal control and the number of internal controls used, thus highlighting the importance of the choice of internal controls in such experiments.
Introduction
Plants have a remarkable ability to cope with highly variable environmental stresses, including cold, drought, and soils with changing salt and nutrient concentrations (i.e. abiotic stress) (Kreps et al., 2002). Nevertheless, these stresses together represent the primary cause of crop loss worldwide (Boyer, 1982), reducing average yields for most major crop plants by more than 50% (Bray et al., 2000). By contrast, the estimated yield loss caused by pathogens is typically around 10–20% (Kreps et al., 2002).
The most important abiotic stress conditions are light, water availability, temperature, and salinity, but also cold and frost can cause severe damage. Plants have developed a variety of defences against low temperatures (Wallis et al., 1997) to which they respond with qualitative and quantitative changes in the observable pattern of gene transcripts and proteins. A number of cDNA clones coding for cold-regulated transcripts and the corresponding genes have been isolated and characterized from a variety of plant species (Cattivelli and Bartels, 1992). Similarly, mechanisms of salt stress have been examined in many plant species. Salt stress involves morphological, physiological, and metabolic changes (Evers et al., 1998).
Concerning biotic stress, plants exposed to virulent and avirulent pathogens respond with a range of defence- and damage-limiting mechanisms. Many of the responses are also induced by other stress-causing agents, which reflects the common elements in different stress situations (Collinge and Boller, 2001). Among biotic stresses in potato, late blight, caused by the oomycete Phytophthora infestans, is one of the most serious diseases of potato and is thus largely studied (Collinge and Boller, 2001; Ghislain et al., 2001; Avrova et al., 2003; Evers et al., 2003). Factors controlling quantitative resistance to P. infestans were found on every potato chromosome, confirming the truly polygenic nature of this trait (Gebhardt and Valkonen, 2001). Although intensively studied, the differences between a resistant and a susceptible response to P. infestans attack are not fully understood (Collinge and Boller, 2001).
Many studies on the defence and stress mechanisms in plants have been based on gene expression (Kirch et al., 1997; Collinge and Boller, 2001; Bezier et al., 2002; Dean et al., 2002). Transcriptome studies have helped to provide a better understanding of plant stress responses. Through these studies, numerous novel stress-responsive genes have been discovered. Overlaps of genes induced by various stress conditions suggest extensive cross-talk between the signalling pathways. Transcriptome studies with multiple time points suggest that plant responses progress from general to specific responses (Sung et al., 2003). The analysis of gene expression requires sensitive, precise, and reproducible measurements for specific mRNA sequences. Gene expression levels were commonly determined using northern blot analysis. However, this technique is time-consuming and requires a large quantity of RNA (Dean et al., 2002). Real-time RT-PCR is, at present, the most sensitive method for the detection of low abundance mRNAs (Bustin, 2000), and can be used for different applications, such as clinical diagnostic (Bustin and Dorudi, 1998), for the analysis of tissue-specific gene expression (Bustin et al., 2000), and for plant studies (Gachon et al., 2004). To avoid bias, RT-PCR is typically referenced to an internal control gene. Ideally, the conditions of the experiment should not influence the expression of this internal control gene (Schmittgen and Zakrajsek, 2000). However, many studies showed that internal standards, mainly housekeeping genes used for the quantification of mRNA expression, could vary with the experimental conditions (Thellin et al., 1999; Warrington et al., 2000; Stürzenbaum and Kille, 2001; Radonic et al., 2004). According to Thellin et al. (1999) and Vandesompele et al. (2002), at least two or three housekeeping genes should be used as internal standards because the use of a single gene for normalization could lead to relatively large errors. Currently, at least nine housekeeping genes are well described for the normalization of expression signals (Stürzenbaum and Kille, 2001). The most common are actin, glyceraldehyde-3-phosphate dehydrogenase, ribosomal genes, cyclophilin, and elongation factor 1-α (ef1α) (Stürzenbaum and Kille, 2001; Bezier et al., 2002; Dean et al., 2002; Thomas et al., 2003). Adenine phosphoribosyl transferase (aprt) (Orsel et al., 2002) and tubulin (Ozturk et al., 2002; Williams et al., 2003) may also be used. Many studies on housekeeping gene expression mainly deal with human tissues (Thellin et al., 1999; Schmittgen and Zakrajsek, 2000; Warrington et al., 2000; Radonic et al., 2004), bacteria and viruses (Stöcher et al., 2002, 2003; Savli et al., 2003), and, as far as is known, only a few have concerned plants, for example, barley (Burton et al., 2004), rice (Kim et al., 2003), poplar (Brunner et al., 2004), Arabidopsis thaliana, and tobacco (Volkov et al., 2003).
In the present work, the variability of expression of seven housekeeping genes (actin, aprt, 18S rRNA, ef1α, tubulin, cyclophilin, and the ribosomal protein L2) and the hsp20.2 gene implicated in late blight stress (Gigliotti et al., 2004), was studied in potato plants exposed to a biotic (late blight) and two abiotic (cold and salt) stresses, in order to assess their value as internal controls in expression studies.
Materials and methods
Plant materials and stress treatments
Quantitative trait loci (QTL) responsible for field resistance to late blight were identified in a cross between the diploid Solanum phureja accession CHS and the S. tuberosum dihaploid clone PS-3. The population is referred as the PD population. The most significant QTL is located on chrXII and contributes up to 43% of the phenotypic variation in some field trials (Ghislain et al., 2001).
Three potato pools, called 00R1, 01R1, and 11R1, formed to represent classes at the targeted QTL of increasing resistance, were used. Plant samples were realized after 0, 1, 2, and 3 weeks of late blight infection in the field (Gigliotti et al., 2004). In order to be able to put as many complete series of samples as possible (11 samples+1 No Template Control) on the same RT-PCR plate experiment, the sample 01R1T2 was omitted from analysis.
Tetraploid Solanum tuberosum ssp. tuberosum L. cv. Desiree was used for cold and salt treatment. Cold treatment of ex vitro potato plants was performed at 4 °C with regard to the control at 20 °C. For salt stress, ex vitro potato plants were treated with 100 mM NaCl, whereas the control was treated with water. For both cold and salt stress, as well as for the control conditions, plant leaves were sampled after 0, 1, 3, 8, and 14 d.
Total RNA extraction
For the late blight experiment, RNA was extracted from plants by the phenol/SDS method (Wilkins and Smart, 1996). RNA was treated with RNase-free DNase I (TaKaRa, Shuzo, Kyoto, Japan).
For cold and salt treatment, RNA was extracted by the RNeasy plant mini kit (Qiagen, Leusden, The Netherlands) including DNase treatment according to the manufacturer's instructions.
Nucleic acid concentrations were measured at 260 nm and were checked by RiboGreen RNA quantitation (Jones et al., 1998) reagent kit according to the manufacturer's instruction (Molecular Probes, Eugene, Oregon, USA) with a TD-700 fluorometer (Turner Designs, Sunnyvale, California, USA). Purity of the total RNA extracted was determined as the 260/280 nm ratio and the integrity was checked by electrophoresis in 1% agarose gel.
Primer design
Seven housekeeping genes were selected: β-tubulin, cyclophilin, actin, elongation factor 1-α (ef1α), 18S rRNA, adenine phosphoribosyl transferase (aprt), and cytoplasmic ribosomal protein L2 gene. The hsp20.2 gene was chosen as the gene of interest. Potato nucleotidic sequences were obtained from the GenBank database (Table 1). For aprt and L2 genes, only a sequence of Arabidopsis thaliana was found. A BLAST against the GenBank EST database permitted the nucleotidic sequences of potato corresponding to aprt and L2 genes to be found (Table 1). The similarity level between Arabidopsis thaliana sequences and the potato EST was 78% on 506 nucleotides and 83% on 774 nucleotides for aprt and L2 genes, respectively.
Primer sequences of seven housekeeping genes and one gene of interest, the amplification length and the melting temperature of the amplified product
Name | Accession number | Primer sequence 5′-3′ | Primer sequence 5′-3′ | Length (bp) | Tm (°C) |
|---|---|---|---|---|---|
| β-tubulin | 609267 | ATGTTCAGGCGCAAGGCTT | TCTGCAACCGGGTCATTCAT | 101 | 79 |
| ef1α | AB061263 | ATTGGAAACGGATATGCTCCA | TCCTTACCTGAACGCCTGTCA | 101 | 79 |
| L2 | 39816659 | GGCGAAATGGGTCGTGTTAT | CATTTCTCTCGCCGAAATCG | 121 | 82 |
| 18S rRNA | X67238 | GGGCATTCGTATTTCATAGTCAGAG | CGGTTCTTGATTAATGAAAACATCCT | 101 | 75 |
| aprt | CK270447 | GAACCGGAGCAGGTGAAGAA | GAAGCAATCCCAGCGATACG | 121 | 80 |
| Actin | X55749 | GCTTCCCGATGGTCAAGTCA | GGATTCCAGCTGCTTCCATTC | 101 | 81 |
| Cyclophilin | AF126551 | CTCTTCGCCGATACCACTCC | TCACACGGTGGAAGGTTGAG | 121 | 81 |
| Hsp20.2 | BQ511516 | TGTTGAAGTTGGGTCTTAGCATAGAAG | CCTCCAGTGCAGGCATGTC | 76 | 78 |
Name | Accession number | Primer sequence 5′-3′ | Primer sequence 5′-3′ | Length (bp) | Tm (°C) |
|---|---|---|---|---|---|
| β-tubulin | 609267 | ATGTTCAGGCGCAAGGCTT | TCTGCAACCGGGTCATTCAT | 101 | 79 |
| ef1α | AB061263 | ATTGGAAACGGATATGCTCCA | TCCTTACCTGAACGCCTGTCA | 101 | 79 |
| L2 | 39816659 | GGCGAAATGGGTCGTGTTAT | CATTTCTCTCGCCGAAATCG | 121 | 82 |
| 18S rRNA | X67238 | GGGCATTCGTATTTCATAGTCAGAG | CGGTTCTTGATTAATGAAAACATCCT | 101 | 75 |
| aprt | CK270447 | GAACCGGAGCAGGTGAAGAA | GAAGCAATCCCAGCGATACG | 121 | 80 |
| Actin | X55749 | GCTTCCCGATGGTCAAGTCA | GGATTCCAGCTGCTTCCATTC | 101 | 81 |
| Cyclophilin | AF126551 | CTCTTCGCCGATACCACTCC | TCACACGGTGGAAGGTTGAG | 121 | 81 |
| Hsp20.2 | BQ511516 | TGTTGAAGTTGGGTCTTAGCATAGAAG | CCTCCAGTGCAGGCATGTC | 76 | 78 |
Primer sequences of seven housekeeping genes and one gene of interest, the amplification length and the melting temperature of the amplified product
Name | Accession number | Primer sequence 5′-3′ | Primer sequence 5′-3′ | Length (bp) | Tm (°C) |
|---|---|---|---|---|---|
| β-tubulin | 609267 | ATGTTCAGGCGCAAGGCTT | TCTGCAACCGGGTCATTCAT | 101 | 79 |
| ef1α | AB061263 | ATTGGAAACGGATATGCTCCA | TCCTTACCTGAACGCCTGTCA | 101 | 79 |
| L2 | 39816659 | GGCGAAATGGGTCGTGTTAT | CATTTCTCTCGCCGAAATCG | 121 | 82 |
| 18S rRNA | X67238 | GGGCATTCGTATTTCATAGTCAGAG | CGGTTCTTGATTAATGAAAACATCCT | 101 | 75 |
| aprt | CK270447 | GAACCGGAGCAGGTGAAGAA | GAAGCAATCCCAGCGATACG | 121 | 80 |
| Actin | X55749 | GCTTCCCGATGGTCAAGTCA | GGATTCCAGCTGCTTCCATTC | 101 | 81 |
| Cyclophilin | AF126551 | CTCTTCGCCGATACCACTCC | TCACACGGTGGAAGGTTGAG | 121 | 81 |
| Hsp20.2 | BQ511516 | TGTTGAAGTTGGGTCTTAGCATAGAAG | CCTCCAGTGCAGGCATGTC | 76 | 78 |
Name | Accession number | Primer sequence 5′-3′ | Primer sequence 5′-3′ | Length (bp) | Tm (°C) |
|---|---|---|---|---|---|
| β-tubulin | 609267 | ATGTTCAGGCGCAAGGCTT | TCTGCAACCGGGTCATTCAT | 101 | 79 |
| ef1α | AB061263 | ATTGGAAACGGATATGCTCCA | TCCTTACCTGAACGCCTGTCA | 101 | 79 |
| L2 | 39816659 | GGCGAAATGGGTCGTGTTAT | CATTTCTCTCGCCGAAATCG | 121 | 82 |
| 18S rRNA | X67238 | GGGCATTCGTATTTCATAGTCAGAG | CGGTTCTTGATTAATGAAAACATCCT | 101 | 75 |
| aprt | CK270447 | GAACCGGAGCAGGTGAAGAA | GAAGCAATCCCAGCGATACG | 121 | 80 |
| Actin | X55749 | GCTTCCCGATGGTCAAGTCA | GGATTCCAGCTGCTTCCATTC | 101 | 81 |
| Cyclophilin | AF126551 | CTCTTCGCCGATACCACTCC | TCACACGGTGGAAGGTTGAG | 121 | 81 |
| Hsp20.2 | BQ511516 | TGTTGAAGTTGGGTCTTAGCATAGAAG | CCTCCAGTGCAGGCATGTC | 76 | 78 |
Eight primer pairs were designed from these sequences (150 bp maximum length, optimal Tm at 60 °C, GC% between 20% and 80%) with the primer express 2.0.0 Applied Biosystems software (Table 1).
Two step real-time RT-PCR
For each gene, PCR efficiency was determined by measuring the CT to a specific threshold (Walker, 2002) for a serial dilution of bulked cDNA. All PCRs displayed efficiencies between 84% and 96%.
Verification of amplified products and sequencing reactions
PCR product sizes were checked on a 4% agarose gel. All corresponded to the expected size. Melting curves showed a single amplified product for all genes and the melting temperatures were in accordance with those calculated (Table 1). In order to verify the sequences of amplification products, PCRs were performed on samples with 300 nM of primers, 2 U of Taq DNA polymerase (Amersham Biosciences, Uppsala, Sweden), 400 μM each of dNTP mix (Takara, Shuzo, Kyoto, Japan), and 10 ng of cDNA in a total volume of 25 μl. Amplifications were performed with the following program: 94 °C 2 min and 40 cycles at 94 °C 30 s, 58 °C 30 s, and 72 °C 30 s. PCR products were purified using the Qiaquick PCR purification kit (Qiagen, Leusden, The Netherlands) according to the manufacturer's instructions. Amplified products were cloned with the TOPO TA Cloning Kit for sequencing (Invitrogen, Merelbeke, Belgium) according to the manufacturer's instructions. Five colonies were analysed by PCR with plasmid specific primers at the final concentration of 200 nM each, T3 (5′-ATTAACCCTCACTAAAGGGA-3′) and M13 (5′-GTAAAACGACGGCCAG-3′) at an annealing temperature of 53 °C. Sequencing reactions were performed in both senses with Big Dye Terminator v3.1 cycle sequencing kit (Applied Biosystems, Applied Biosystems, Foster City, USA). 100 nM of primers cited above, 1 μl of PCR product, 2 μl of Big Dye Mix, and 2 μl of sequencing buffer were used in a total volume of 25 μl. Sequence reactions were run on the ABI PRISM 310 Genetic Analyzer (Applied Biosystems, Applied Biosystems, Foster City, USA).
Sequences of amplification products were compared to GenBank sequences used to design primers with BLAST 2 sequences software (http://www.ncbi.nlm.nih.gov/blast/bl2seq/bl2.html). All amplified sequences had 100% identities with GenBank sequences.
Data acquisition
Expression levels were determined as the number of cycles needed for the amplification to reach a threshold fixed in the exponential phase of PCR reaction (CT) (Walker, 2002). The CT were transformed into quantities using PCR efficiencies according to Vandesompele et al. (2002) in order to use geNorm software (http://allserv.ugent.be/∼jvdesomp/genorm/index.html).
Statistical analysis
CT values from the ABI PRISM 7000 Sequence Detection System (Applied Biosystems, Foster City, USA) were analysed. A two sample F-test was performed in order to compare two population variances. A P-value superior to 0.05 indicated that no difference of variation of expression could be deduced.
Results
Variations of housekeeping gene
To evaluate the stability of expression of housekeeping genes, RNA transcription levels for all samples were measured for each stress. The RNA transcription profiles of the seven housekeeping genes and the gene of interest are shown (Fig. 1). 18S rRNA showed the highest expression level (low CT value). The CT values were similar for the three stresses (Fig. 1). Whatever the stress was, the genes appeared organized in the same order according to their level of expression. The RNA transcription level varied among stresses and, as could be shown by a F-test (P >0.05), actin was the most variable gene for late blight and salt stress. No significant variation was found for cold stress. In order to choose the best housekeeping genes, geNorm analysis (Vandesompele et al., 2002) was used. Vandesompele et al. (2002) defined two parameters to quantify the housekeeping gene stability: M (average expression stability) and V (pairwise variation). A low M value is indicative of a more stable expression, hence, increasing the suitability of a particular gene as a control gene. Vandesompele et al. (2002) proposed 0.15 as a cut-off value for the pairwise variation below which the inclusion of an additional control gene is not required. Depending on the stress, the more stable housekeeping genes were not the same ones (Fig. 2). For late blight exposure, the most stable genes were ef1α and 18S rRNA. The M value obtained for these two genes was 0.217 and the V value was 0.070 (Fig. 2), so there was no need to add a third gene as an internal control. The two most stable genes for cold stress were ef1α and L2, and ef1α and cyclophilin for salt stress. The M values were 0.408 and 0.305 for cold and salt stress, respectively. As for late blight exposure, salt stress had a pairwise variation below the cut-off (0.119), so only two genes need to be used as an internal control. For cold stress, the pairwise variation was higher than 0.15 (0.165), so a third gene should be added in order to normalize gene expression. The least stable genes during abiotic stresses were actin and tubulin as they significantly increased the pairwise variation for both stresses as shown in Fig. 2d by increasing V values. The most stable gene present in the three stresses was ef1α.
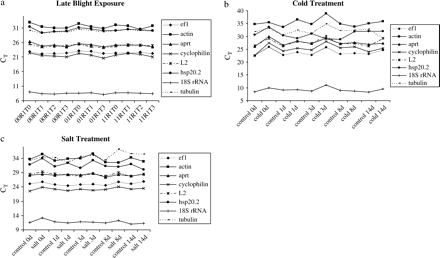
RNA transcription levels of housekeeping genes tested, presented as CT mean value in the different samples. (a) Genotypes 00R1, 01R1, and 11R1 exposed to late blight for 0, 1, 2, or 3 weeks. (b) Solanum tuberosum cv. Desiree exposed to cold treatment for 0, 1, 3, 8, or 14 d. (c) Solanum tuberosum cv. Desiree exposed to salt treatment for 0, 1, 3, 8, or 14 d. CT values are mean of three replicates.
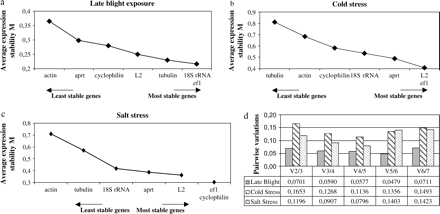
Average expression stability values of control genes by geNorm analysis: (a) late blight exposure, (b) cold treatment, (c) salt treatment, and (d) determination of the optimal number of control genes for normalization by geNorm analysis.
hsp20.2 expression
The expression level for the gene of interest was quantified according to geNorm instructions (Vandesompele et al., 2002). The hsp20.2 gene expression increased after 1 week of late blight exposure for the genotype 00R1 and remained constant after 2 and 3 weeks of exposure (Fig. 3a). For the other genotypes, the level of hsp20.2 expression remained constant whatever the time of exposure. In the genotype 00R1 the expression level was 2.5-fold higher after 1 week of exposure; this genotype was the most sensitive to late blight among the genotypes used in this study (Fig. 3a). During cold stress, hsp20.2 expression increased after 1 d at 4 °C. At 3 d of treatment, the gene expression level was the highest and increased 15.6-fold compared with the control. After 3 d, the level of expression decreased so as to reach its initial expression level (Fig. 3b). During salt stress, the expression level increased until day 14 of treatment and the maximum expression level was obtained after 14 d of exposure. The increase was 4-fold at 14 d with regard to the control (Fig. 3c).
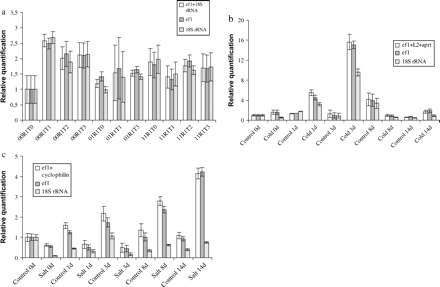
Relative quantification of hsp20.2 expression using a unique internal control (18S rRNA or ef1α) and with two or three of the most stable housekeeping genes defined by the geNorm analysis: ef1α+18S rRNA for late blight, elfα+L2+aprt for cold stress, and ef1α+cyclophilin for salt stress (a) during late blight stress, (b) during cold stress, and (c) during salt stress.
Many authors use 18S rRNA to quantify gene expression level as a unique internal control. Thus the expression level of the hsp20.2 gene was quantified with one internal control (18S rRNA or ef1α) and with two or three of the most stable housekeeping genes as shown previously by geNorm: 18S rRNA+ef1α for late blight, ef1α+cyclophilin for salt stress and ef1α+L2+aprt for cold stress. Differences in quantification were detected according to the internal controls used. Compared with the use of 18S rRNA, using ef1α as the internal control did not significantly change the quantification of the expression level of hsp20.2 (Fig. 3). However, when 18S rRNA was used as a unique internal control, the quantification of the expression was underestimated for both salt and cold treatment (Fig. 3).
Discussion
For relative RT-PCR to be accurate, specific PCR conditions and an appropriate internal control must be determined. A reliable internal control should show minimal changes, whereas a gene of interest may change greatly over the course of an experiment (Dean et al., 2002). Thus, choosing an internal control is very important to quantify gene expression. As shown (Fig. 3), differences in quantification were observed depending on the number and the choice of housekeeping genes. The majority of studies in the literature use a unique internal control. The actin gene was often used to normalize the quantification of expression (Bezier et al., 2002; Langer et al., 2002; Thomas et al., 2003). However, in this study, this gene did not appear to be the best gene to use as some variations of expression during the different treatments appeared. The 18S and 28S ribosomal subunits are other examples of commonly used internal controls (Burleigh, 2001; Gonzalez et al., 2002; Klok et al., 2002) that are subject to controversy. Indeed, when directly compared to other housekeeping genes, their expression level was extremely stable (Stürzenbaum and Kille, 2001). Thellin et al. (1999) recommended the use of 28S or 18S rRNA as internal standards for mRNA quantification studies because mRNA variations were weak and could not highly modify the total RNA level. There are several arguments against the use of rRNA as the internal control. Ribosomal subunit transcription is affected by biological factors and drugs (Vandesompele et al., 2002). Further drawbacks to the use of 18S or 28S rRNA molecules as standards are their absence in purified mRNA samples, and their high abundance compared with target mRNA transcripts. The latter makes it difficult to subtract the baseline value in real-time RT-PCR data analysis accurately (Vandesompele et al., 2002). As ribosomal subunits are not polyadenylated they cannot be exploited when dealing with cDNA derived from total RNA utilizing oligo-dT primers in the RT reaction. It is precisely for this reason that the ribosomal subunits have failed to replace the use of other housekeeping genes (Stürzenbaum and Kille, 2001). For the hsp20.2 gene, relative quantification varied depending on the internal control used. When 18S rRNA was used in both cold and salt stresses, the variations of hsp20.2 expressions in the samples were weaker compared with the use of ef1α.
hsp20.2 gene expression during late blight exposure normalized with 18S rRNA as the internal control showed similar results to those obtained with ef1α or with the two housekeeping genes used. In this experiment, 18S rRNA belonged to the two best housekeeping genes found with geNorm software as opposed to the other stress experiments. The use of 18S rRNA as the internal standard could be a valuable alternative to quantify genes of interest, keeping in mind that it could reduce the variations of expression. The quantification of expression of hsp20.2 using ef1α as the single housekeeping gene led to similar results as those obtained by using two housekeeping genes. These results were in accordance with Dean et al. (2002) and with Stürzenbaum and Kille (2001) who stipulated that elongation factor-1 α was a good invariant control. The use of several housekeeping genes (at least two) was recommended by Thellin et al. (1999) in order to compare gene expression levels to housekeeping gene transcripts as internal standards. According to Vandesompele et al. (2002) the purpose of normalization was to remove the sampling difference (such as RNA quantity and quality) in order to identify real gene-specific variation. They provided evidence that a conventional normalization strategy based on a single gene led to erroneous normalization.
Several studies showed that Hsps play a crucial role in protecting plants against abiotic stresses (Sabehat et al., 1996, 1998; Visioli et al., 1997; Wang et al., 2004). Among the five conserved families of Hsps (Hsp70, Hsp60, Hsp90, Hsp100, and the small Hsp), the small Hsp are the most prevalent in plants (Visioli et al., 1997; Wang et al., 2004). In this study Hsp20.2, a small Hsp was implicated in salt and in cold stress with a maximum of expression at 14 d and 3 d, respectively. The results of cold stress were similar to those shown by Van Berkel et al. (1994) where a cDNA clone corresponding to a small heat shock protein (22.3 kDa) had maximum induction after 3 d of cold storage and decreased during prolonged cold storage. They proposed that a transient expression pattern might be an indication that the gene product functions in the process of cold adaptation. Many mechanisms explaining the phenomenon of cross-tolerance suggested that specific proteins were induced in several kinds of stress (Sabehat et al., 1998). Avrova et al. (2003) showed the implication, at an early stage, of several heat shock proteins upon Phytophthora infestans infection, but no small Hsp was then found. In the material used this study, hsp20.2 was previously described to be expressed in potato infected by Phytophthora infestans (Gigliotti et al., 2004). An increase of hsp20.2 expression was observed in the most sensitive genotype (00R1) whereas the level of expression of hsp 20.2 remained constant for the other genotypes. Thus this study indicates that small heat shock proteins might be implicated in late blight resistance.
To conclude, for the quantification of hsp20.2 expression, the choice of internal standards is very important to normalize the level of expression. In this study ef1α was the only housekeeping gene tested that was usable for normalization in the three experiments. The expression of elongation factor 1-α did not seem to be influenced during cold, salt, and late blight stresses and it could thus be used to normalize expression levels of genes of interest.
The authors gratefully acknowledge L Solinhac for technical assistance, M Ferreol for his statistical support and Dr N Kieffer (University of Luxembourg) for the use of the RT-PCR machine. They thank the International Potato Center (Lima, Peru) and especially M Ghislain, for providing the potato RNA samples of the plants infected by late blight.
References
Avrova AO, Venter E, Birch PRJ, Whisson SC.
Bezier A, Lambert B, Baillieul F.
Bray EA, Bailey-Serres J, Weretilnyk E.
Brunner AM, Yakovlev IA, Strauss SH.
Burleigh SH.
Burton RA, Shirley NJ, King BJ, Harvey AJ, Fincher GB.
Bustin SA.
Bustin SA, Dorudi S.
Bustin SA, Gyselman VG, Siddiqi S, Dorudi S.
Cattivelli L, Bartels D.
Collinge M, Boller T.
Dean JD, Goodwin PH, Hsiang T.
Evers D, Ghislain M, Hausman JF, Dommes J.
Evers D, Hemmer K, Hausman JF.
Gachon C, Mingam A, Charrier B.
Gebhardt C, Valkonen JPT.
Ghislain M, Trognitz B, Herrera MR, Solis J, Casallo G, Vàsquez C, Hurtado O, Castillo R, Portal L, Orrillo M.
Gigliotti S, Hausman JF, Evers D.
Gonzalez MC, Echevarria C, Vidal J, Cejudo FJ.
Jones LJ, Yue ST, Cheung CY, Singer VL.
Kim BR, Nam HY, Kim SU, Kim SI, Chang YJ.
Kirch HH, Van Berkel J, Glaczinski H, Salamini F, Gebhardt C.
Klok EJ, Wilson LW, Wilson D, Chapman SC, Ewing RM, Somerville SC, Peacock WJ, Dolferus R, Dennis ES.
Kreps JA, Wu Y, Chang HS, Zhu T, Wang X, Harper JF.
Langer K, Ache P, Geiger D, Stinzing A, Arend M, Wind C, Regan S, Fromm J, Hedrich, R.
Orsel M, Krapp A, Daniel-Vedele F.
Ozturk ZN, Talamé V, Deyholos M, Michalowski CB, Galbraith DW, Gozukirmizi N, Tuberosa R, Bohnert HJ.
Pfaffl MW.
Radonic A, Thulke S, Mackay IM, Landt O, Siegert W, Nitsche A.
Sabehat A, Weiss D, Lurie S.
Sabehat A, Weiss D, Lurie S.
Savli H, Karadenizli A, Kolayli F, Gundes S, Ozbek U, Vahaboglu H.
Schmittgen T, Zakrajsek BA.
Stöcher M, Leb V, Berg J.
Stöcher M, Leb V, Hölzl G, Berg J.
Stürzenbaum SR, Kille P.
Sung DY, Kaplan F, Lee KJ, Guy CL.
Thellin O, Zorzi W, Lakaye B, De Borman B, Coumans B, Hennen G, Grisar T, Igout A, Heinen E.
Thomas C, Meyer D, Wolff M, Himber C, Alioua M, Steinmetz A.
Van Berkel J, Salamini F, Gebhardt C.
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F.
Visioli G, Maestri E, Marmiroli N.
Volkov RA, Panchuk II, Schöffl F.
Wallis JG, Wang H, Guerra DJ.
Wang W, Vinocur B, Shoseyov O, Altman A.
Warrington JA, Nair A, Mahadevappa M, Tsyganskaya M.
Wilkins T, Smart LB.

{kind=link}
{kind=link}
{kind=link}
Comments